Algen sind die am einfachsten organisierten Pflanzen. Viele von ihnen sind einzellig, einige noch ohne Zellwand, andere mit Zellwänden, die sich in ihrer Zusammensetzung und Struktur deutlich von denen höherer Pflanzen abheben. Bei ihnen können wir die Evolution der Zellwand zurückverfolgen. Primitive Zellwände erfüllen nicht die für höhere Pflanzen geltenden Ansprüche.
Eine Struktur wie die Zellwand ist im Verlauf der Evolution der Organismen zweifelsfrei mehrfach entstanden. Alle Archaebakterien, Eubakterien, und mit ihnen die Blaualgen (Cyanophyta oder Cyanobacteria) besitzen komplex gebaute Wände, deren Biosynthese energetisch recht aufwendig ist. Weder was die Zusammensetzung noch was den Biosynthesemodus angeht, haben sie irgendwelche Gemeinsamkeiten mit pflanzlichen Zellwänden.
Obwohl die Abstammung der Pflanzen aus frühen eukaryotischen Zellen im Detail ungeklärt ist, besteht Konsens darüber, daß die primitiven Algen Flagellaten sind und den übrigen, nicht grünen Flagellaten nahestehen. Viele, doch nicht alle Arten auf dieser Organisationsstufe, zu denen als typische grüne Vertreter die Euglenophyta gehören, sind zellwandlos. Von der Umwelt sind sie jedoch nicht nur durch eine einfache Membran getrennt, sondern durch eine bereits recht komplex organisierte Pellikula. Sie besteht im wesentlichen aus Glykoproteinen, die in regelmäßigen Mustern in der Art eines zweidimensionalen Kristalls organisiert sind. In regelmäßigen Intervallen ist die Zelloberfläche von helical angeordneten Rippen und dazwischenliegenden Furchen umwunden.
Echte Zellwände findet man bei den meist einzelligen Algen, so auch bei den Volvocales. Die am eingehendsten untersuchte Art ist Chlamydomonas reinhardii. In ihrer Wand fehlen langkettige fibrilläre Kohlenhydrate. Glykoproteine machen die Hauptmasse aus, wobei bereits hier ein dem Extensin ähnliches hydroxyprolinreiches Protein auftritt. Zu den nachgewiesenen Zuckerresten gehören Arabinosyl-, Galactosyl- und Mannosylreste. Elektronenmikroskopisch scheint die Wand aus sieben Schichten aufgebaut zu sein. Die mittlere enthält ein ausgedehntes gitterförmiges Raster aus polygonalen Platten, die vornehmlich aus dem genannten Glykoprotein bestehen, während die darüber- bzw. darunterliegenden Ansätze faserförmiger Strukturen zeigen. Die äußerste variiert in ihrer Mächtigkeit, denn sie ist aus Komponenten aufgebaut, die die Zelle der Umgebung entnimmt.
Damit wird bereits eine wesentliche Aufgabe der Zellwand einfacher, einzelliger Arten deutlich: Sie ist Mittler zwischen Zelle und Umwelt, dient also nicht nur ihrem Schutz sondern zugleich auch der Kommunikation mit gieichartigen und andersartigen Zellen. Sie muß für Metaboliten und Regulatormoleküle durchlässig sein und/oder Rezeptormoleküle tragen, mit deren Hilfe eine Zelle Kontakt zu einer anderen aufnehmen kann. Aus der Vielfalt dieser Aufgaben (und Spezifitäten) ergibt sich die Evolution einer Vielfalt unterschiedlich strukturierter Zellwände.
In pflanzlichen Vielzellern ist die Kommunikation entlang der gesamten Zelloberfläche weitgehend eingeschränkt. Nachbarschaftsbeziehungen der Zellen entwickeln sich im Verlauf der Entwicklung von Geweben. Festigkeit ist ein entscheidendes Kriterium. Stoffaustausch zwischen Zellen erfolgt über spezifische Öffnungen in der Wand (Tüpfel, Plasmodesmen). Die ursprünglich von einer Einheit geleisteten Funktionen sind somit auf zwei funktionelle Strukturen aufgeteilt.
Die wesentlichen Strukturelemente aller pflanzlichen Zellen sind Polysaccharide, deren unterschiedliche chemische Zusammensetzung grundsätzlich verschiedene physikalische Eigenschaften nach sich zieht. Es gibt keine pflanzliche Zellwand, die nur aus einer Molekülklasse besteht( Die Wechselwirkungen zwischen den verschiedenen Molekülen führt zu Merkmalen, durch die sich Zellwände bestimmter Klassen voneinander unterscheiden.
Cellulose ist schon bei vielen Algenklassen das Hauptstrukturelement der Wand. Die Fibrillenstruktur hingegen zeigt noch beträchtliche Variationen. Es gibt sichere röntgenstrukturanalytische Beweise dafür, daß sie auch in Algenzellwänden - zumindest über weite Strecken - kristallin ist. Unterschiede in der Art der Beugungsreflexe (ein Maß für periodische Abstände im molekularen Bereich), die von Art zu Art beträchtlich schwanken (besonders groß bei den Rhodophyta [Rotalgen]), weisen darauf hin, daß die Cellulose in vielen mehr oder weniger gleichförmigen kristallinen Formen zusammengelagert sein kann.
Bei einigen Klassen der Algen gibt es lediglich Streutexturen, bei anderen (vor allem bei vielen Arten der Chlorophyta) liegt ein höherer Organisationsgrad (Schichten parallel gelagerter Mikrofibrillen) vor. Solche Schichten wechseln jedoch in der Regel mit Schichten amorphen Materials ab. Bei den meisten Algen entfällt die klare Unterscheidung von Primär- und Sekundärwand, und wo sie existiert, beruht deren Existenz auf einem andersartigen Bildungsmodus als bei den höheren grünen Pflanzen.
Mannane: Bei einer Reihe mariner Grünalgen (Codium, Dasycladus, Acetabularia u.a.), und in der Wand einiger Rotalgen (Porphyra, Bangia), bilden Mannane die Hauptstrukturelemente. Auch sie sind linear, und die Mannosylreste sind beta 1 > 4 glykosidisch miteinander verknüpft. Wasserstoffbrücken können gebildet werden; sie sind (wie bei der Cellulose) auch die Ursache für die partiell kristalline Organisation in Mikrofibrillen. Bei Codium sind sie fest mit dem Protein assoziiert.
Xylane: Polymere, in denen beta-D-Xylosylreste über 1 > 3 und 1 > 4 glykosidische Bindungen verknüpft sind. Sie sind als Strukturelemente in einigen Rot- und Grünalgen nachgewiesen worden. Im Gegensatz zu den bisher besprochenen Polymeren haben wir es hier - zum Teil wenigstens - mit verzweigten Strukturen zu tun; dennoch findet man bei Arten, deren Wand Xylane enthält, einen schichtigen Aufbau und eine Orientierung von Mikrofilamenten. In ihnen sind lineare Polymere vorherrschend.
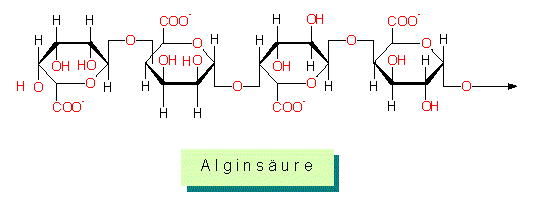
Alginsäure: Alginsäure und ihre Salze, die Alginate sind wichtige Bestandteile der Wände von Phaeophyta (Braunalgen). Sie sind in vielerlei Hinsicht einzigartig. Sie bestehen ausschließlich aus Uronsäuren: Mannuronsäure und beta-L-Glucuronsäure in wechselnden Verhältnissen, und in geringeren Mengen auch aus beta-D-Glucuronsäure.
Homopolymere kommen neben Heteropolymesen vor, zum Teil findet man artspezifische Unterschiede, was wiederum ein Hinweis darauf ist, daß die einzelnen Arten mit einem unterschiedlichen Sortiment an Enzymen ausgestattet sind.
Die Alginate der Braunalgen kommen sowohl in als auch außerhalb der Zellwand (in der Interzellularsubstanz) vor. Ihr Anteil an der Zellwand kann bis zu 40 Prozent der Trockenmasse ausmachen. Sie haben eine hohe Affinität zu divalenten Kationen (Calcium, Strontium. Barium, Magnesium) und die Eigenschaft, zu gelieren. Die Hauptmenge der aus Braunalgen isolierbaren Magnesium - Ionen entstammt der Alginsäurefraktion.
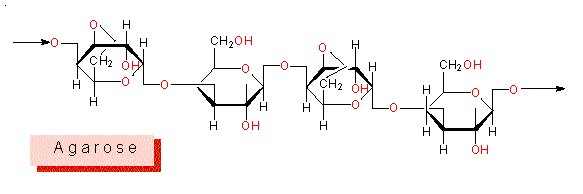
Sulfonierte Polysaccharide: Polysaccharide, deren Monomere mit Sulfonsäureresten verestert und darüber hinaus zum Teil methyliert sind, wurden bei nahezu allen marinen Algen nachgewiesen. Sie kommen zum Teil in der Zellwand selbst, teilweise in der Interzellularsubstanz vor. Sulfonierte Galactane sind für viele Rotalgen typisch, herkunftsgemäß werden sie als Agarose, Carrageenan, Porphyran, Furcelleran und Funoran bezeichnet.
Sowohl L- als auch D-Galactose, verknüpft durch beta 1 > 3 oder alpha 1 > 4 glykosidische Bindungen, bilden das Grundmuster der Agarose und des Porphyrans (dort: alternierend L- und D-Galactosylreste), während Carrageenan und Furcelleran ausschließlich die D-Form enthalten. Wie bei den Alginaten, ist Gallertbildung eine der wichtigsten physikalischen Eigenschaften dieser Molekülfamilie. Agar, dessen Grundeinheit Agarose ist, wird vornehmlich aus den Rotalgengattungen Gelidium und Gracillaria gewonnen.
Die in Agarose und Carrageenan vorkommenden außergewöhnlichen Bindungstypen führen zu spezifischen Tertiärstukturen.
Weitere Zellwandbestandteile. Eine Reihe von Algen enthalten mineralische Zellwandkomponenten. So ist das Silizium als wesentlicher Bestandteil der Diatomeenschalen bekannt, doch tritt es auch in Zellwänden anderer Algengruppen auf, so ist z.B. der Chrysophyt Synura von siliziumhaltigen Schuppen umgeben, bei einigen Braunalgen und bei der Grünalge Hydrodictyon ist es ein Bestandteil der Wand. Diatomeen nehmen Silizium als Silikat auf. Der Vorgang ist sauerstoff- und temperaturabhängig, energieverbrauchend und auf die Anwesenheit divalenten Schwefels angewiesen.
Sporopollenin ist ein Isoprenderivat. Es ist Bestandteil von Pollenzellwänden, doch darüber hinaus wurde es in Wänden einiger Grünalgen (Chlorella, Scenedesmus u.a.) nachgewiesen.
Calcium: Calciumeinlagerungen in Zellwände sind verschiedentlich beschrieben worden. Besonders häufig scheinen sie bei Arten tropischer, mariner Gewässer zu sein. Einige der Arten sind an Riffbildungen beteiligt. Calcium wird durchweg als Calciumcarbonat abgelagert, von dem (mindestens) zwei unterschiedliche kristalline Formen vorkommen: Calcit (wird gebildet in Wänden einiger Gruppen der Rotalgen und Charophyceen) und Argonit [gebildet durch einige Grünalgen (Acetabularia u.a.), Braun- und Rotalgen]. Gemische beider Formen in einer Art kommen nicht vor.
|
|