
Seit Beginn der experimentellen Pflanzenphysiologie befaßte man sich mit dem Einfluß von Licht auf die Entwicklung der Pflanzen. Hervorzuheben sind die von J. v. SACHS entwickelten Versuchseinrichtungen und die späteren Arbeiten von W. PFEFFER und J. BONNER.
Im Dunkeln kultivierte Keimlinge zeichnen sich in der Regel durch intensives Streckungswachstum aus. Die Internodien rücken unverhältnismäßig weit auseinander, Blattanlagen werden gebildet, doch eine Ausdifferenzierung zu Blattspreiten unterbleibt. Die Sprosse sind gelblich, da fast kein Chlorophyll entsteht. Vorsicht Ausnahmen: Tradescantia albiflora, dann einige Gymnospermen, und viele niedere Pflanzen bilden auch bei Dunkelheit Chlorophyll. Die durch Lichtmangel gestörte Entwicklung wird Etiolement genannt, die Pflanzen sind etioliert.
Ein Überangebot an Licht, z.B. eine starke UV-Strahlung im Hochgebirge, führt zu drastisch reduziertem Streckungswachstum der Internodien, zu einer Verkleinerung der assimilierenden Oberfläche, vielfach zu starker Anthocyanbildung und meist zu intensiver gefärbten Blüten.
Die hier skizzierten, bereits im vorigen Jahrhundert bekannten Befunde wurden in den letzten Jahrzehnten durch Aussagen aus gezielten Experimenten ergänzt und gestützt. Hierbei ging es zunächst einmal um die Frage, welche der Differenzierungs- und Wachstumsleistungen lichtabhängig sind, wie das Wirkungsspektrum aussieht und welche Lichtmengen benötigt werden.
H. MOHR und Mitarbeiter (Institut für Biologie, Universität
Freiburg) haben durch Untersuchungen an Sinapis alba-Keimlingen
eine Vielzahl lichtinduzierter Reaktionen analysiert. Im Gegensatz zu vielen
niederen (vorwiegend aquatischen) Pflanzen, bei denen lichtinduzierte Reaktionen
meist, aber nicht ausschließlich, durch kurzwellige Strahlung (Blaulicht)
ausgelöst werden, ist bei der Photomorphogenese der höheren Landpflanzen
in der Regel eine langwelligere Strahlung ausschlaggebend. Hierauf aufbauend,
entwickelten H. A. BORTHWICK, S. B. HENDRICKS und Mitarbeiter (Plant Industry
Station, US Department of Agriculture, Beltsville, Md.) im Zeitabschnitt
zwischen 1946 und 1959 das Konzept des Hellrot/Dunkelrot-, bzw. Phytochromsystems.
|
Einige durch Licht bewirkte Reaktionen des Senfkeimlings (Sinapis
alba).
|
|
Hemmung des Hypokotyl-Längenwachstums |
 Das
Phytochrom ist ein Chromoprotein, dessen Zustand durch Licht beeinflußt
wird. Es wird vorwiegend bei Dunkelheit gebildet und liegt zunächst
als PR vor (P ist die Abkürzung für Phytochrom, R
steht für red). Nach Bestrahlung mit lambda= 660 nm Licht (hellrot,
red) geht es in PFR über [FR=far red; auf deutsch
könnte man auch schreiben: PHR (hellrot), resp. PDR
(dunkelrot); oder unter Verwendung der Wellenlängen P660,
resp P730]. PFR wird durch lambda = 730 nm
Licht (dunkelrot) in PR rücküberführt. PR
ist die biologisch inaktive, PFR die biologisch aktive Form.
Das
Phytochrom ist ein Chromoprotein, dessen Zustand durch Licht beeinflußt
wird. Es wird vorwiegend bei Dunkelheit gebildet und liegt zunächst
als PR vor (P ist die Abkürzung für Phytochrom, R
steht für red). Nach Bestrahlung mit lambda= 660 nm Licht (hellrot,
red) geht es in PFR über [FR=far red; auf deutsch
könnte man auch schreiben: PHR (hellrot), resp. PDR
(dunkelrot); oder unter Verwendung der Wellenlängen P660,
resp P730]. PFR wird durch lambda = 730 nm
Licht (dunkelrot) in PR rücküberführt. PR
ist die biologisch inaktive, PFR die biologisch aktive Form.
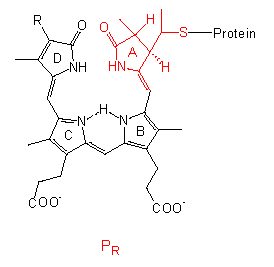
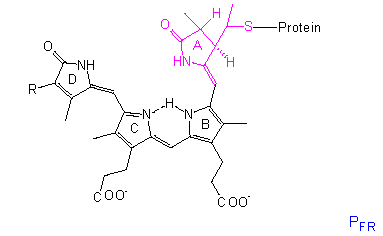
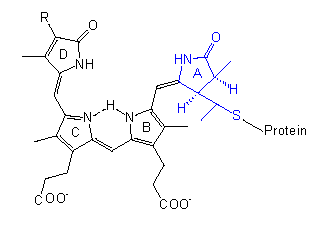
Die chromophore Gruppe ist ein lineares Tetrapyrrol, dessen Konformation und Absorptionsspektrum sich in der PR-Form deutlich von der in PFR unterscheidet. Eine ähnliche Gruppe mit vergleichbaren Konformationsänderungen liegt in den Bilirubinen der Rotalgen vor, dort steht lediglich anstelle der Vinylgruppe am Ring D eine Äthylgruppe.
Das Protein ist ein Dimer, es besteht aus gleichen Untereinheiten, deren Molekulargewichte je nach Pflanzenart zwischen 120000 und 127000 liegen; folglich ist es ein allosterisches Protein. Nach allem, was wir aus der Proteinstrukturforschung wissen, wirkt eine Strukturänderung im Chromophor wie ein Hebel und zieht eine Konformationsänderung des Proteins nach sich; der allosterische Effekt bedingt eine weitere Amplifikation (Signalverstärkung). Dadurch ändern sich natürlich auch die Bindungseigenschaften des Phytochroms zu anderen Molekülen, so daß eine Kaskade in Gang kommen kann, an deren Ende die meßbaren physiologischen Phänomene stehen. Ob das Phytochrom direkt als Effektor fungiert oder ob weitere Moleküle zwischengeschaltet sind, bleibt zu klären. Vielleicht ist das auch von Fall zu Fall verschieden.
Für eine Reihe von Genprodukten ließ sich zeigen, daß das Phytochrom auf Transkriptionsebene (mRNS) kontrollierend in das Geschehen eingreift. Das gilt u.a. für die kerncodierte kleine Untereinheit der Ribulose-1,5-Bisphosphatcarboxylase und das Chlorophyll - a / b-bindende Protein. Darüber hinaus wurde gezeigt, daß es in Form einer Rückkopplungsschleife die Expression seines eigenen Gens reguliert.
PR wird bei Dunkelheit im Cytoplasma gebildet und dort so lange akkumuliert, bis ein bestimmter Spiegel erreicht ist. Es stellt sich ein Gleichgewicht zwischen Synthese und (langsamem) Abbau ein. Die Überführung in PFR nach Bestrahlung mit hellrotem Licht ist ein rasch ablaufender Prozeß. PFR ist außerordentlich instabil, der Phytochromspiegel in der Zelle sinkt daher nach Belichtung auf ein bis drei Prozent des ursprünglichen Werts, der vermutlich ein neues Gleichgewicht zwischen PR-Synthese und PFR-Abbau repräsentiert. Nach Verdunklung steigt die Phytochrommenge erneut aufgrund einer de novo-Synthese von PR. Die PFR-Eliminierung beruht also nicht allein auf einer Proteininaktivierung, sondern auf einer zusätzlichen Inaktivierung translatierbarer mRNS. Die negative Rückkopplung wird nach Verdunklung aufgehoben, da der PFR-Spiegel auf nahezu Null absinkt; gleichzeitig steigt die mRNS-Menge wieder an (P. H. QUAIL und Mitarbeiter, University of Wisconsin, Madison).
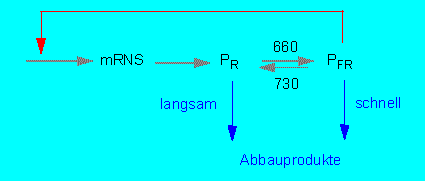
Gleichgewichtslage und Regulationsvermögen des Phytochromsystems (Nach P. H. QUAIL, 1984)
Weitere Einzelheiten über den Wirkungsmechanismus erhielt man durch den Einsatz gentechnischer Verfahren (die mRNS wurde in cDNS überschrieben und kloniert, Nukleotidsequenzen phytochrom-codierender Gene sind bekannt) sowie monoklonaler Antikörper gegen bestimmte Domänen des Proteins. Selbst Arabidopsis thaliana, eine Pflanze mit einem der kleinsten Genome enthält 5 Phytochromgene, die Nukleotidsequenzen der einzelnen Gene variieren beträchtlich. Diese und Daten, die an anderen Arten ermittelt wurden, weisen darauf hin, daß Phytochrome einer sehr variablen Proteinfamilie mit mindestens 3 Unterfamilien (phyA, phyB, phyC) angehören, die bereits vor der Abgliederung der Monokotyledonen von ihren Dikotyledonenvorfahren entstanden sind. Aufrund der proteinanalytischen Daten lassen sich zwei generell unterschiedliche Photorezeptortypen ausmachen: Typ I oder Phytochrom etiolierter Gewebe und Typ II, das Phytochrom grüner Gewebe. In etiolierten Geweben kommt Typ I in großen Mengen vor, unterliegt aber einem hohen Turnover, Typ II findet sich dort in kleinen Mengen im stabilen Zustand PFR. Da das Phytochrom Typ I von phyA Genen codiert wird, bezeichnet man es nun auch als Phytochrom A, das Typ II - Phytochrom hingegen ist recht heterogen, es besteht aus mindestens 2 immunologisch nicht verwandten Polypeptidketten (die auch mit dem Phytochrom A keine immunologische Verwandtschaft aufweisen). Die Polypeptidketten der Phytochrome haben Längen von 1110-1172 Aminosäureresten und gehören damit zu den längsten Polypeptiden überhaupt; jede Polypeptidkette trägt einen Chromophor und faltet sich zu zwei Domänen. Es sieht nunmehr so aus, als könnte die Heterogenität der Phytochrome einerseits, bedingt durch gleichartige chromophore Gruppe, auf das gleiche Lichtsignal reagieren, aufgrund der unterschiedlichen Proteinstrukturen das Signal dann aber an unterschiedliche Rezeptoren weiterleiten und somit eine Anzahl physiologisch unterschiedlicher Reaktionen induzieren (QUAIL, 1991).
Nachdem gezeigt war, daß das Phytochrom in die Transkriptionskontrolle eingreift, stellte sich die Frage, ob es auf DNS-Ebene spezifische Erkennungsregionen gibt, die eine lichtabhängige Transkription des entsprechenden DNS-Abschnitts bewirken. G. MORELLI et al. (Rockefeller University, New York, 1985) zeigten - wiederum unter Einsatz gentechnischer Verfahren -, daß ein 33 Basenpaare langes Stück, welches die TATA-Box (Teil des Promoters) einschließt und das dem Gen für die kleine Untereinheit der Ribulose-1,5-Bisphosphatcarboxylase vorgeschaltet ist, für die lichtinduzierte Steuerung der Genexpression essentiell ist. Inzwischen wurde die Lichtabhängigkeit der Transkription für eine ganze Anzahl von Proteinen nachgewiesen, dazu gehören das Chlorophyll a/b bindende Protein, die a - Untereinheit der ATP Synthetase, das 32-kDa Protein des Photosystem II, die Chalcon-Synthetase u.a. (SIMPSON und HERRERA-ESTRELLA, 1990), auf der DNS wurden je nach Gen sowohl upstream, als auch downstream (also am Anfang einer Transkriptionseinheit und an ihrem Ende) Nukleotidsequenzen ermittelt, die für die lichtinduzierte Steuerung benötigt werden und die damit für die Modulation der mRNS-Synthese, d.h. für die Menge der gebildeten mRNS-Sequenzen (Transkripte) verantwortlich sind (GILMARTIN et al., 1990).
Durch indirekte Immunfluoreszenz ließ sich Phytochrom im Plasma der Zellen, im Zellkern und den Plastiden lokalisieren. Es ist nicht in allen Zellen in gleicher Menge enthalten. Man findet man es z.B. in der Epidermis ausschließlich in den Schließzellen.
Das Phytochrom ist an der Induktion der Chloroplastendrehung der fädigen Grünalge Mougeotia beteiligt. Man unterscheidet zwischen der Schwachlicht- und der Starklichtstellung des plattenförmigen Chloroplasten (Flächenstellung, Kantenstellung). Wir haben es hier also mit einer intrazellulären Bewegung zu tun, denn die Lichtperzeption erfolgt in jeder Zelle getrennt. Es gibt demnach zumindest auf dieser Ebene auch keinen Informationsaustausch zwischen benachbarten Zellen. Selbst innerhalb einer Zelle ist die Bewegung der einzelnen Chloroplastenabschnitte autonom. Nachweisbar ist der Effekt durch eine partielle Bestrahlung einer Zelle.
W. HAUPT (Botanisches Institut der Universität Erlangen, 1970) benutzte Mikrostrahlen polarisierten Lichts, mit denen eine Zelle Punkt für Punkt abgetastet werden konnte. Dadurch ließ sich beweisen, daß das den Lichtreiz perzipierende Phytochrom hier an der Peripherie der Zelle (aller Voraussicht nach im Plasmalemma) liegt und in einer bestimmten Weise orientiert ist.
Der Chloroplast reagiert auf eine Bildung (Anreicherung) von PFR . Bei dessen Bildung aus PR drehen sich die Photorezeptoren um 90 Grad. PR ist parallel zur Zelloberfläche orientiert, PFR senkrecht dazu. Der Chloroplast dreht sich daher stets von Orten höchster PFR-Konzentration weg.
Bliebe zu fragen, wodurch die Drehung selbst bewirkt wird. Es sieht so aus, als sei das Aktin daran beteiligt; doch wie das Lichtsignal in Bewegungsenergie umgesetzt wird und wie die Aktinfilamente an Chloroplasten ansetzen, bleibt nach wie vor offen.
Eine orientierte Ausrichtung von Phytochrommolekülen an der Zellperipherie ist nicht auf Mougeotia beschränkt. Eine ähnliche Erscheinung liegt der phototropen Reaktion von Farnchloronemen zugrunde. Andererseits ist Phytochrom - vor allem bei Algen - nicht das dominierende Sensorpigment. Blaulicht hat einen stärkeren Einfluß; mit steigender Wassertiefe nimmt der Anteil kurzwelligen Lichts bekanntlich zu. Das auf langwelliges Licht adaptierte Phytochromsystem hätte keinerlei Vorteile.
Für die Lichtperzeption bei der Chloroplastenbewegung verschiedener Algen und anderer Organismen, sind vier physikalisch unterschiedliche Konzepte erkannt worden.
Es gibt eine Reihe physiologischer Prozesse, beispielsweise die Anthocyanbildung, die erst bei länger andauernder Bestrahlung aktiviert oder auf maximale Leistung gebracht werden. Das Wirkungsspektrum umfaßt einen weiten Lichtwellenbereich; mehr als man aufgrund der Absorptionsspektren des Phytochroms erwarten dürfte. Offensichtlich wird demnach mehr als nur ein Sensorpigment aktiviert.
K. M. HARTMANN (seinerzeit Institut für Biologie, Universität Freiburg) zeigte bereits 1966, daß das Wachstum von Salatkeimlingen durch getrennt gegebenes Licht der Wellenlängen lambda = 658 nm und lambda = 768 nm unwirksam ist, während eine simultane Bestrahlung mit Licht dieser beiden Wellenlängen zu einer Wachstumszunahme führt. Alles weist darauf hin, daß auch diese Reaktionen allein über Phytochrom laufen. Man benötigt nur die Zusatzannahme, daß es außer PR und PFR weitere Formen: PRX, PFRX, PRX' und PFRX' gibt, die reversibel (direkt oder indirekt) ineinander übergehen und in der Zelle in einem Gleichgewicht zueinander stehen. Es sieht also so aus, als gäbe es einen Energietransfer zwischen den Sensorpigmenten (oder deren unterschiedlichen Aktivitätszuständen), wodurch die Empfindlichkeit gegenüber Licht stark moduliert werden kann. Das Signal erfährt dadurch eine Verstärkung (Amplifikation). Das Wirkungsspektrum der Anthocyanbiosynthese weist auf die Existenz weiterer Pigmente hin. Viel spricht für zusätzliche Photorezeptoren für den Blau- und UV-Anteil des Spektrums. Solche sind in der Tat nachgewiesen worden (Cryptochrom [Blaulichtrezeptor] und der UV-B-Photorezeptor), und es konnte gezeigt werden, daß beide modulierend auf das Phytochromsystem einwirken. Für die Pflanze wäre es von Vorteil, weil sie damit exakt und unmittelbar auf die an natürlichen Standorten herrschenden Lichtbedingungen reagieren könnte. Bei Algen stimuliert Blaulicht die Carotinoidsynthese, die Chlorophyllsynthese und den Glucoseabbau; bei einigen marinen Arten wird auch die Thylakoidbildung beeinflußt. Bei Acetabularia wirkt es als Zeitgeber der endogenen Rhythmik.
|
|