
Das Zusammenklappen der Fiederblättchen und Blätter einiger Pflanzenarten (vor allem aus der Gruppe der Leguminosen und der Gattung Oxalis) nach Berührung oder Erschütterung, gehört seit dem Altertum zu den faszinierendsten Erscheinungen in der Botanik. Man bezeichnet solche Pflanzen als sensitiv. Vergleichbare Reaktionen findet man auch beim Zusammenklappen der Blatthälften der Venusfliegenfalle (Dionaea muscipula), den Tentakelbewegungen der zur gleichen Familie gehörenden Drosera-Arten sowie bei den Staubfäden der Centaurea- und Berberis-Arten und der Bewegung der Narbenhälften von Mimulus.
Schon früh suchte man nach Erklärungen für diese Erscheinungen. Wie auch bei allen anderen bisher besprochenen Fällen, müssen wir die Bewegung einerseits und die Reizwahrnehmung, -weiterleitung und -umsetzung andererseits getrennt betrachten. Es lag nahe, die Bewegung der Mimosenblätter mit den tagesperiodischen Bewegungen mancher Blätter zu vergleichen, allein schon deshalb, weil sie bei den Leguminosen besonders eindrucksvoll zu beobachten sind.
R. H. J. DUTROCHET (1776-1847, Mediziner und Privatgelehrter in der Touraine und in Paris) vermutete (1837), daß das Heben und Senken der Blätter durch einen Antagonismus der oberen und unteren Gelenkhälften hervorgerufen wird. Direkte Messungen durch den Physiologen E. W. von BRÜCKE (1848) ergaben, daß der Turgor in den Blattgelenken zunimmt, wenn Blätter die Schlafstellung einnehmen, wobei die Zunahme auf die Gelenkoberseite beschränkt bleibt. Damit war gesichert, daß es sich tatsächlich um Turgorbewegungen handelt. Durch die ergänzenden Untersuchungen von J. v. SACHS (1859) und W. PFEFFER (1873) wurde die Aussage erhärtet.
Pflanzenanatomische Untersuchungen ergaben, daß die Biegefestigkeit im Gelenkbereich reduziert ist, weil die sonst im Stengel und in Blattstielen peripher liegenden Leitbündel (= Festigungsgewebe) zu einem zentral gelegenen Strang vereint sind, der von großzelligen Rindenparenchymzellen umgeben ist. Sie sind dazu prädestiniert, ihren Turgor durch reversible Wasseraufnahme zu ändern. Die hohe Elastizität ihrer Wände bietet eine weitere Voraussetzung für Formveränderungen (Deformationen). Die Zellen der Gelenkoberseite sind in der Lage, ihren Turgor weit über das in Zellen normale Maß zu steigern; die Zellen der Unterseite haben die Eigenschaft, überdurchschnittlich leicht Wasser zu verlieren und sind daher besonders stark deformierbar.
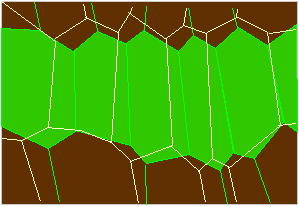
Schema des Ziehharmonikagewebes in der Unterseite des Primärgelenks von Phaseolus-Zellen voll turgeszent (grün) oder erschlafft (in Umrissen). (Nach W. HAUPT, 1977).
Ein geregelter Bewegungsablauf setzt ein gut aufeinander abgestimmtes Zusammenspiel der beiden antagonistischen Gelenkhälften voraus. Daß das tatsächlich der Fall ist, haben bereits die von v. BRÜCKE angestellten Messungen ergeben.
Die Situation im Gelenkbereich zwischen den beiden Blatthälften von Dionaea muscipula sieht ähnlich aus. Jede der Blatthälften trägt auf ihrer Oberseite je drei Sinnesborsten, und nur über sie erfolgt die Reizung. Die Betrachtung ihres anatomischen Baus legt die Annahme nahe, daß hier eine Hebelwirkung vorliegt, denn auch an der Basis der Borsten liegt eine Gruppe großer deformierbarer Zellen.
Daß wir es mit nastischen Bewegungen zu tun haben, geht aus der Anatomie der Bewegungsgewebe hervor (dorsiventraler Bau). Wegen der Reaktion auf Erschütterung spricht man von Seismonastie. Reaktionen, die auf einen Berührungsreiz hin erfolgen, werden im allgemeinen als Thigmonastien bezeichnet. Die Seismonastie ist zweifelsohne nur als ein Grenz- oder Spezialfall der Thigmonastie zu betrachten, der sich durch eine besonders hohe Geschwindigkeit der Reizumsetzung auszeichnet. Bei der Erschütterung oder Berührung einzelner Fiederblättchen der Mimose klappen sie paarweise zusammen. Die Reaktion beginnt an der Berührungsstelle und breitet sich von dort aus sukzessive über das gesamte Fiederblatt aus, überträgt sich dann auf das nächstliegende Gelenk, von dort aus auf das nächste usw. Dabei lassen sich zwei wichtige Feststellungen machen:
- Die Reaktion verläuft nach einer Alles-oder-Nichts-Regel.
- Sie ist reversibel. Nach 15 bis 30 Minuten kehren die Blätter in ihre Ausgangsstellung zurück.
Es gibt also auf jeden Fall eine Erregungsleitung im Blatt, und damit stellt sich für uns die Frage nach ihrem Mechanismus. Reaktionen dieser Art kennt man von tierischen Nervenzellen (Neuronen) und weiß, daß die Erregungsfortleitung entlang des Axons einer Nervenzelle auf elektrochemischem Wege erfolgt. Mit anderen Worten: die Membran wird lokal depolarisiert, und die Depolarisierung überträgt sich mit hoher Geschwindigkeit auf benachbarte Bereiche. Der Aufbau des Axons gewährleistet eine gerichtete Ausbreitung. Spielen solche Membrandepolarisierungen auch bei der Mimose (und bei reizbaren Organen anderer Arten: Dionaea, Drosera, Berberis usw.) eine Rolle? Es gibt gesicherte Anhaltspunkte dafür, daß das tatsächlich der Fall ist, obwohl es experimentell außerordentlich schwierig ist, Änderungen des Membranpotentials pflanzlicher Zellen zu bestimmen. Aus Messungen an Zellen anderer Arten, vor allem der Riesenzellen der Alge Nitella, aber auch von Zellen des Bewegungsgewebes der Mimose, lassen sich folgende Aussagen machen:
Weil die Konzentrationen bestimmter Ionen aufgrund unterschiedlicher selektiver Permeabilität der Membran inner- und außerhalb der Zelle verschieden sind, bildet sich ein meßbares Membranpotential aus, z. B., weil Kalium - Ionen entlang ihres Konzentrationsgefälles von innen nach außen diffundieren und somit eine Ladungstrennung über der Membran herbeiführen und ihre Gegenionen (meist Chlorid - Ionen) die Membran gar nicht oder nur in geringem Ausmaß passieren können. Gegenüber der Gesamtionenkonzentration ist die Menge der über die Membran verschobenen Ionen außerordentlich gering. Die Ruhepotentiale betragen bei Pflanzen -150 bis -200 mV (Nitella, Elodea, Vallisneria). Während eines Aktionspotentials erhöht sich die Permeabilität der Membran für Chlorid - Ionen, die dann entsprechend ihres Konzentrationsgradienten ausströmen und die Membran somit depolarisieren. Diese Depolarisation erhöht nun ihrerseits den Ausstrok von Kalium - Ionen (Repolarisationsphase). Diese erfolgt nicht durch eine aktive Aufnahme von Ionen,
Eine Reizung der Zelle (wie auch immer) führt zu einem vorübergehenden lokalen Zusammenbruch des Potentials, der auf kurzfristiger Ionendurchlässigkeit der Membran beruht. Das Ausströmen von Kationen bewirkt einen vorübergehenden Anstieg des Potentials (Aktionspotential), das unmittelbar nach dem Durchlaufen seines Maximums wieder zum alten Zustand zurückkehrt. Der Zeitraum bis zum Erreichen des Ruhepotentialwerts ist die Refraktärzeit. Während dieser Zeitspanne ist jener Membranbereich nicht reizbar. Eine Depolarisation kann sich leicht und vor allem sehr schnell (im msec-Bereich) auf die übrigen Membranabschnitte der Zelle ausbreiten und damit die Gesamtmembran der Zelle erfassen. Wenn das Aktionspotential einer Zelle ausreichend hoch ist, kann es Nachbarzellen erregen. Voraussetzung ist allein das Überspringen der Reizschwellen jener Zellen. Das erklärt einmal eine Reizweiterleitung von Zelle zu Zelle, und zum anderen die Alles-oder-Nichts-Regel.
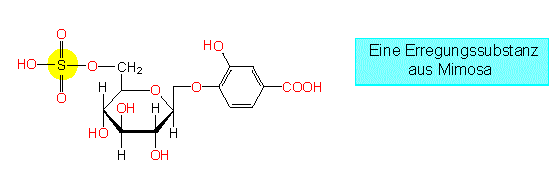
Zellen im Bewegungsgewebe einer Mimose lassen sich außer auf mechanischem Wege auch durch einen elektrischen Stromstoß reizen, ein Hinweis darauf, daß eine elektrische Reizleitung existiert. Zusätzlich konnte die Freisetzung einer Substanz aus gereizten Zellen nachgewiesen werden, die durch den Transpirationsstrom verbreitet wird, auf diese Weise ebenfalls die Gelenke erreicht und sie zu einer Reaktion veranlaßt.
Elektrische und chemische Reizung oder Erregungsleitung unterscheiden sich in ihren Geschwindigkeiten voneinander. Je nach Organ beträgt sie 0,7-26 cm/sec für die elektrische, und 0,15-2 cm/sec für die chemische Reizleitung. Beide Arten ergänzen einander. Die chemische Reizleitung ist überall dort von Vorteil, wo elektrische Widerstände auftreten, z.B. an Gelenken und in toten Zellbereichen.
|
|