
Der See hat das Profil eines Kegels, der Durchmesser beträgt maximal 400 Meter, die Oberfläche 14 Hektar und das Volumen 1,3 Millionen Kubikmeter. Er erreicht eine Tiefe von 29,5 Metern. Da er von einem geschlossenen Waldgürtel umgeben ist, ist er weitgehend windgeschützt. Es gibt keine Zu- und Abflüsse. Er wird als eutroph eingestuft.
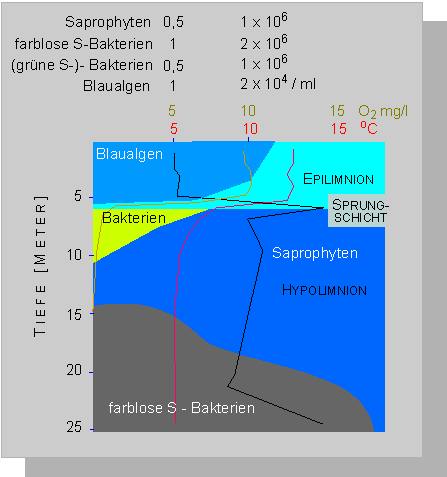
Gliederung des Plußsees (Ostholstein) am 7. 10. 1964
(J. OVERBECK, 1972)
Das Epilimnion hat eine Mächtigkeit von fünf Metern, die Sprungschicht ist in den Sommermonaten deutlich ausgeprägt. Im Phytoplankton dominieren die Blaualgen in wechselnder Artenzusammensetzung. Oscillatoria redekii, Anabaena flos-aquae und Aphanizomenon gracile sind in aufeinanderfolgenden Jahren in unterschiedlichen Anteilen vertreten. Es besteht kein Zusammenhang zwischen der Algenpopulation im Plußsee und der in einem der benachbarten Seen. So ist unter anderem die genannte Aphanizomenon-Art außer im Plußsee in keinem der anderen Seen der ostholsteinischen Seenplatte in größerer Zahl enthalten. Welche der Arten in einem bestimmten Jahr dominiert, ist schwer vorhersagbar; mit entscheidend sind offensichtlich die Startbedingungen zu Saisonbeginn, d.h., die Anzahl teilungsfähiger Zellen zu einem Zeitpunkt, der für eine Massenvermehrung optimal ist.
Das saisonale und vertikale Verteilungsprofil von Oscillatoria redekii in den Jahren 1964 bis 1967 gemessen worden.
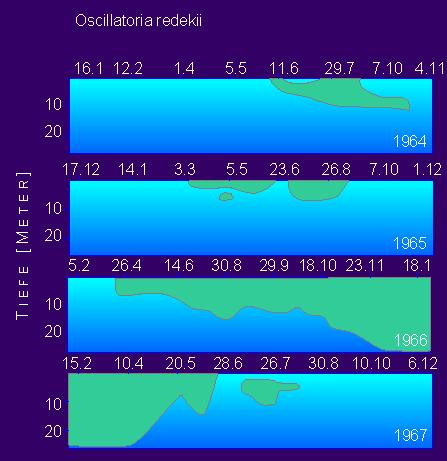
Vorkommen von Oscillatoria redekii (grüne Flächen) im Plußsee in den Jahren 1964-1967 in einer 25 m tiefen Wassersäule
(Nach J. Overbeck, 1972
Auffallend ist das nur sporadische Auftreten in den Jahren 1964 und 1965; 1966 war die Art den ganzen Sommer und Herbst über dominierend. Mit abnehmender Wassertemperatur sank die Sprungschicht stetig ab, die Grenze zwischen Epi- und Hypolimnion löste sich auf; im gesamten Wasserkörper des Sees erfolgte eine Vollzirkulation, die zu einer Durchmischung des Wassers mit Nährstoffen und Sauerstoff führte. Die Folge war eine den gesamten Wasserkörper umfassende Ausbreitung der Oscillatoria im Winter 1966/67.
Die Anzahl der planktischen Algenarten ist - von Wintermonaten mit Eisgang abgesehen - über das ganze Jahr hinweg mehr oder weniger konstant. Die Biomasse des Phytoplanktons hingegen durchläuft in jeder Saison ein, zwei oder mehr deutlich ausgeprägte Maxima. Das Frühjahrsmaximum wird in der Regel durch massenhaftes Auftreten von Diatomeen und Peridineen hervorgerufen. Sie erreichen hohe Zellzahlen bei niedriger Wassertemperatur. Es folgt (Messungen aus dem Jahr 1979) ein Flagellatenmaximum (Eudorina, Cryptomonas u.a.); das Sommermaximum wird durch Oscillatoria redekii gebildet, und im Herbst, nach dem Verschwinden der Blaualgen, trat Rhizochysis scherfelii (eine Chrysophycee) in großer Zahl auf.
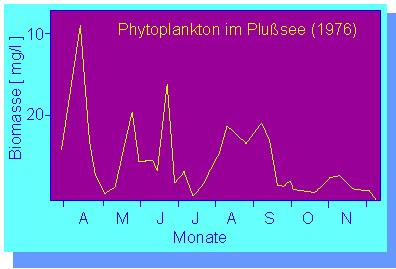
Saisonale Verteilung der Phytoplanktonbiomasse. Einzelheiten über die Artzusammensetzung s. Text
(Nach B. HICKEL, 1977)
Eine stets wiederkehrende Frage lautet, warum eine bestimmte Algenpopulation so plötzlich die Überhand gewinnt, aber nach kurzer Wasserblüte genauso plötzlich wieder in sich zusammenbricht. Eine vielzitierte Hypothese der Phytoplanktonökologie besagt, daß die interspezifische Konkurrenz um Nährstoffe die Artenzusammensetzung und die saisonbedingte Sukzession des Phytoplanktons in Seen determiniert.
Der Aufbau einer Population ist recht einfach erklärbar. Zu Beginn der warmen Jahreszeit ist der See nährstoffreich, die Lichtwerte erreichen für das Wachstum optimale Werte. Problematischer sind die Erklärungen für den Abfall der Populationsgröße. Langjährige Messungen ergaben, daß der Plußsee phosphatlimitiert ist. Zwar ist anorganisches Phosphat im Hypolimnion stets in ausreichender Menge vorhanden, im Epilimnion hingegen sinken die Werte bereits im Mai/Juni so stark ab, daß in dem betreffenden Jahr nicht mehr mit einer übermäßigen Biomasseproduktion zu rechnen ist. Das anorganische Phosphat wird im Verlauf einer Saison vom Phytoplankton restlos aufgebraucht. Es kann dann bis in den September/Dezember hinein dauern, bevor sich der Pool wieder auffüllt. Der Phosphatmangel ist sicherlich der gravierendste Faktor für den Zusammenbruch einer Algenblüte.
Ein weiterer Faktor, der - zumindest zeitweilig - eine Dominanz der Blaualgen sichert, ist deren Tendenz, Antibiotika und andere toxische Substanzen ins Medium abzugeben, und damit konkurrierende Arten zu unterdrücken. Manche Blaualgen sezernieren eisen-chelierende Agentien und entziehen damit durch selektive Entfernung eines essentiellen Ions den übrigen Arten die Nahrungsgrundlage (T. A. MURPHY et al., 1976).
Phytoplankton ist ein Glied in der Nahrungskette, und ein reichhaltiges Angebot ist die Grundlage einer starken Zooplanktonvermehrung. Als Begleitorganismen sind oftmals Pilze (Chytridien) nachgewiesen worden, und gegen Ende einer Algenblüte war kaum ein Filament pilzfrei, so daß anzunehmen ist, daß der Pilzbefall den Zusammenbruch der Algenblüte beschleunigt. Ein großer Teil des absterbenden Phytoplanktons sinkt ab und sammelt sich im Sediment. Der bakterielle Abbau ist die Ursache der Sauerstoffzehrung im Hypolimnion (Aktivität Methan-oxydierender Bakterien).
Parallel dazu erfolgt eine bakterielle Sulfatreduktion, bei der große Mengen an Schwefelwasserstoff anfallen. Der so entstehende Schefelwasserstoff-haltige Lebensraum erstreckt sich gegen Ende des Sommers nahezu über den gesamten Tiefenbereich des Sees und begünstigt das Auftreten roter und grüner phototropher Schwefelbakterien. Ihre ökologische Bedeutung liegt im Aufbau organischer Substanz unter Ausnutzung von Restlicht im anaeroben Bereich, wodurch das Schwefelwasserstoff zur Kohlendioxyd-Reduktion verbraucht wird. Die Pigmentzusammensetzung der Bakterien erlaubt es ihnen, die Restlichtmenge in größerer Tiefe zu nutzen. Für 1980 konnten im Plußsee die folgenden Eckdaten des Kohlenstoffkreislaufs ermittelt werden:
1. Primärproduktion: 25 000 kg C* / Jahr [* C = Kohlenstoff]
2. Methanproduktion: 7400 kg C / Jahr,
3. Methanoxydation: 6075 kg C / Jahr.
Daraus folgt, daß 82 Prozent des im See gebildeten Methans im Verlauf eines Jahres oxydiert werden. Die Relation der Methanproduktion zur Primärproduktion (Photosynthese der Phytoplankter) liegt bei 29 Prozent. Der Wert ist für eutrophe Seen typisch. Geht man nunmehr davon aus, daß etwa 10 Prozent der Primärproduktion (hier also 2500 kg C / Jahr) den Seeboden erreichen und daß davon 85 Prozent (1875 kg C / Jahr) zu Methan abgebaut werden, verbleibt ein Fehlbetrag von 5525 kg C / Jahr, der in Form anderer Verbindungen dem See zugeführt werden muß (Niederschläge, Erosion usw.). In solchen Einträgen ist das gemessene Kohlenstoff-Phosphor-Verhältnis ca. 100:1; d.h., es müssen außer den 5525 kg C / Jahr auch etwa 50 kg Phosphor pro Jahr in den See gelangen. Dieser Wert liegt im Bereich, der zusätzlich erforderlich ist, um die Menge der gebildeten Biomasse zu erklären.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de