
Atoms, ions and molecules are constantly moving, colliding and thus changing their direction. In an ideal gas move the single particles independently of one another. The particle distribution is not influenced by these movements and no predominant or preferred direction exists. The velocity depends on the temperature and increases when the temperature rises. It is therefore also spoken of thermal movements (T). The particle movements within liquids are more restricted but the movements within a solution are subject to the same criteria as those in an ideal gas. Often can the plasma of a cell, too, be regarded as such a solution. Although the description does not fit for all cases (just think of the compartmentalisation).
If a concentrated solution of any substance like, for example, sugar, is layered with water do the layers at first stay clearly separated. Since we assumed that dissolved particles (in our case sugar molecules) move in a strictly statistical manner, will at first single particles cross from one to the other liquid and their number will increase with time [f (t)]. Some of them, though not all will return and a net flow of particles moving in the direction formerly free of sugar molecules will develop.
Particle movements from high to lower concentrations are called diffusion. Only after an even distribution is achieved can no further net movements be observed, the system has reached an equilibrium.
How can this process be described quantitatively? The number of particles that cover a certain distance by diffusion is dependent on the cross section's area of the chosen vessel (F) and a substance constant (D).
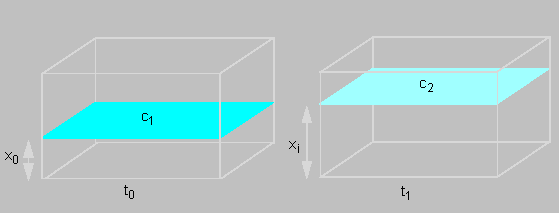
The net flow can now be described as a function of time:
f(t) = DF (c1 - c2 / x1 - x0)
c1 and c2 are the concentrations of the chosen planes, x1 - x0 is the distance between them. Since the concentration decreases with growing distance has the concentration gradient a negative value. If the formula is applied to any small distance then the values have to be given as differentials:
dm / dt = DF (dc / dx ) [cm2 sec-1]
The precept expressed in this formula is also called FICK's first law of diffusion. It states that a substance diffuses in the direction that eliminates its concentration gradient dc / dx at a rate proportional to the magnitude of its gradient.
The dimensions of the quantities are m: [Mol], F: [square centimetre], c: [Mol/ cubic centimetre], x: [cm].
The number of moles that passes a certain plane per second is called the net flow (Phi):
Phi = - D (dc / dx) [Mol cm-2 sec-1]
or
Phi = - D (cl - c2 / x1 - x0)
or, when eliminating D:
D = -Phi / (cl - c2 / x1 - x0)
(c1 - c2/ x1 - x0) is called the concentration gradient. D is the diffusion constant that is defined as the amount of a substance (in Mol) that diffuses through a certain area (in square centimetre) at a concentration gradient of 1 (Mol/ cm).
Diffusion is very quick over short distances but extremely slow at long ones. It is the square of the distance that has an influence on the formula and it is proportional to the available area. Diffusion is important when regarding the molecular movements within cells or between neighbouring cells. But it looses its significance when regarding whole tissues and ceases to have any effect at all when, for example, thinking of the assimilate transport from leaves to roots.
Permeability is the term that describes the diffusion of particles through membranes. It is unimportant whether we regard natural or artificial membranes (like plastic film). We will in a rough approximation assume that the membrane equals a solvent layer of the thickness d in order to obtain a quantitative statement.
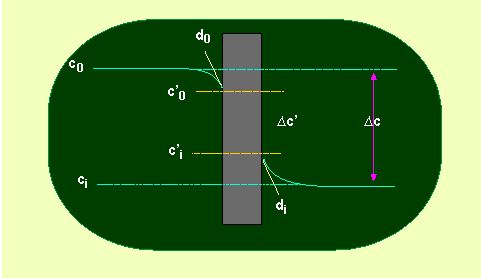
From this assumption is the flux through a membrane directly proportional to the diffusion constant. The constant is without exception lower than in water. The net flux through the membrane can therefore be described as:
Phi = - D (c1 - c2 / d) = - D / d (c1 - c2)
D / d is the permeability constant and has the dimension [cm/ sec].
Biological membranes are not equally permeable for all substances. They are selectively permeable, i.e. the membranes can be permeated by a substance A but not by a substance B. Strictly speaking are they only permeable for some gases like oxygen, nitrogen, carbon dioxide, etc., because only these can permeate a membrane by free diffusion. As has been shown above, is diffusion strictly subject to the laws of thermodynamics. Molecules are characterized by specific properties like size, ability to be converted into a ion or solubility within a membrane. Such lipophilic (fat soluble) molecules can cross the membrane more easily than hydrophilic ones.
Membranes contain a number of further integral components of which proteins are most the important in this context. By a specific arrangement of membrane components can pores or channels be formed that can be selectively permeated by certain ions and molecules. Their permeability is on one hand dependent on the diameter and on the other hand on the electric charge of the pore. Small anions like chloride can easily pass positively charged pores. Cations are detained. Such a state is also called a restricted diffusion. Numerous small molecules like sugar and amino acids seem to be preferentially transported through membranes. But their permeation kinetics does not follow the formula mentioned above, it resembles more an enzyme - substrate turnover kinetic. Such a kinetic is known as facilitated diffusion. The permeability is selective and dependent on the presence of specific membrane-bound transport molecules with an affinity to a selected group of chemically related substances. The substrates do not pass the membrane by themselves but are transported as a substrate-carrier complex. In summary is facilitated diffusion characterized as follows:
- The membrane contains specific transport or carrier molecules.
The carriers (mostly proteins, but also some antibiotics like valinomycin) bind to the substrate (sugar or amino acid, for example). In contrast to enzymes do they not convert their substrates but transport them through the membrane instead.
Several substrates can compete for the same carrier. Their binding properties concerning the carrier may differ and as a consequence differ their transport kinetics, too.
The loaded carrier crosses the membrane with an other velocity than the unloaded carrier.
Die treibende Kraft der erleichterten Diffusion ist wie bei einfacher Diffusion
The maximal transport velocity is dependent on the number of the carrier molecules in the membrane.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de