
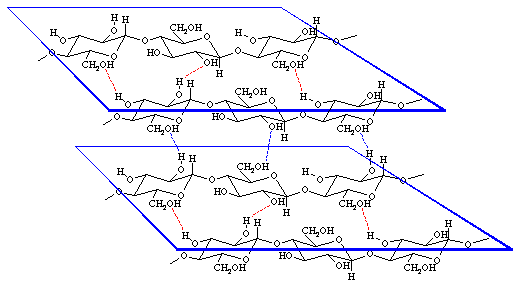
Cellulose is composed of linear chains of covalently linked glucose residues. It is very stable chemically and extremely insoluble. In the primary cell wall consists one glucose polymer of roughly 6000 glucose units, in the secondary wall is their number increased to 13 - 16000 units. Cellulose chains form crystalline structures called microfibrils. A microfibril with a diameter of 20 - 30 nm contains about 2000 molecules.
Crystalline and non-crystalline sections alternate. In crystalline ones forms the cellulose three-dimensional lattices due to the formation of the highest possible number of hydrogen bonds. This high degree of organization is not achieved in the other sections, called paracrystalline. Crystals polarize light. By studying cellulose between crossed polarisators can the main orientation of the microfibrils be determined. In the primary wall do they occur in every possible orientation (disperse texture). During the development of the secondary wall are they deposited in layers (as lamellas). The microfibrills of each layer are parallel to each other (parallel texture). Their orientation changes from layer to layer. Often, especially in very strong cell walls (like those of cotton) are the microfibrills arranged screw-like around the cell's axis. In such cases changes the turning angle from layer to layer (screw-like texture).
Although cellulose is by far the most common macromolecule - nature synthesizes roughly 1011 tons per year and breaks most of it down again - is astonishingly little known about its biosynthesis. The enzyme (or the enzyme complex?) cellulose synthase is still a largely hypothetical quantity.
Since the preparation techniques (freeze etching) were optimized several years ago have directed particles (or particle complexes) been observed at the outer side of the plasma membrane. It is assumed that they take part in the synthesis of cellulose. In some algae (Micrasterias, Spirogyra and also in cells of higher plants) do the complexes form hexagonal rosettes (R. M. BROWN, D. MONTEZINOS, 1976; O. KIERMAYER and U. B. SLEYR, 1979; W. HERTH, 1983).
A striking correlation of the orientation of cortical microtubuli and the neighbouring microfibrills (spatially separated by the plasma membrane) exists. Despite of existing criticism does it look as if the microtubuli would take part in the orientation of cellulose synthase (directed arrangement of the complexes seen in the electron microscope) and would thus exert an indirect influence on the orientation of the microfibrills. Experiments with microtubuli-disrupting agents have an influence on the orientation pattern of newly developing microfibrills.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de