

Fig. 17: Formation of a net-like structure. Differentiated cells (blue squares) form a trace behind moving activator maxima (red). Differentiated cells remove a substrate (green wavy lines). Since the activator/inhibitor system depends on this substrate, the activator maxima are shifted to that neighbouring cells that have the largest distance from other differentiated cells. This is usually the cell at the tip of the filament. The patterning process comes to rest if a certain density of the filaments is reached.
Filamentous branching structures are a common pattern element in all higher organisms. They are used to supply the tissue with nutrition, water, oxygen and information. The venation of leaves, the tracheae of insects, the blood or lymph vessels as well as neurones are examples. How can such complex patterns emerge?
According to the model proposed (Meinhardt, 1976, 1981), such a structure results if local signals are generated that cause an elongation of the filaments. The latter repell the signal, causing in this way its further shift of the signal and thus its further elongation. In the simulation Fig. 17, a local high activator concentration is used as the signal to cause stable activation of a gene when a threshold is exceeded. The exposed cells differentiate and become, for instance, a part of a vascular system. To orient the elongation into a region not supplied by filaments, the filaments repel the signal (red). This occurs if the activator-inhibitor production depends on the concentration of a substrate that is produced by all cells but removed by the differentiated cells. The substrate concentration (green in Fig. 17) is a measure of how urgently the cells need the ingrowth of veins, for instance, in order to remove an oxygen deficiency. The signal (red) will be shifted into a neighbouring cell which will differentiate and become thus a part of the vascular system, too. This is usually the cell at the tip of a filamemt. A repetition of this process - differentiation, shift of the signal, differentiation - leads to a long strand of differentiated cells behind a wandering activator maximum (Meinhardt, 1976, 1982, 1998). For tracheae in insects, all the predicted ingredients have been found (see Shilo et al. 1997; Hacohen et al., 1998).
Branches are formed whenever activator maxima become sufficiently remote from each other during elongation of filaments. Then, the inhibitor concentration can become locally so low that a new activator maxima is triggered along an existing vein due to a small baseline activator production of the differentiated cells. Whether a branch is formed towards the one side or the other depends on minute fluctuations. However, if a branch has been formed, let us say, to the right, the next branch will probably point to the left due to the substrate removal by the first branch. Such alternation in the branching direction can be seen in many leaves. If the details of a pattern depend on fluctuations but each step has a strong influence on forthcoming decisions, the actual pattern will be unpredictable. Indeed, the venation of two leaves on the same tree are never identical although they are certainly formed under control of the same positional information. With increasing density of the filaments the further elongation can be switched off whenever a certain density is obtained.
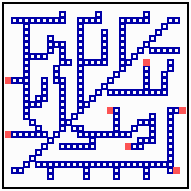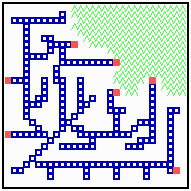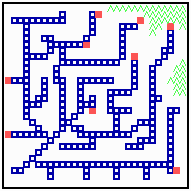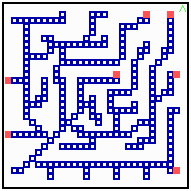
Fig. 18: Regeneration in a net-like structure. After removal of some filaments, the substrate increases in that area , new filaments grow in and the defect becomes repaired.
After removal of some filaments, the system is able to regenerate the missing veins (or whatever it is) since in these regions, the substrate is no longer removed and the rising substrate concentrations attract activator maxima from the non-injured region (Fig. 18). The regenerated pattern is similar but not identical. In nerves and tracheae, the elongation of the filaments does not occur by accretion of new differentiated cells but by local elongation of single cells.