|
|
|
|
|
GENETICS |
|
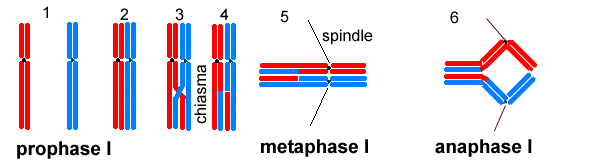
Mendelian inheritance At the beginning of meiosis, in prophase I the nuclear envelope disintegrates and chromosomes become visible as in mitosis (1). The chromosomes have replicated but individual chromatids are not visible. Instead of lining up on a metaphase plate, as in mitosis, chromosomes come together in pairs (2). Each chromosome in a pair is similar in structure (homologous), but would have come originally from different parents. Later in prophase the homologous pairs twist round each other and chromatids may cross over (3). Breaks occur at these cross-overs (or chiasmata, singular chiasma) and pieces of chromatid are exchanged (4). Chromosome pairs line up across the equator of the spindle at metaphase I (5). In anaphase I the chromosomes separate and travel to opposite ends of the spindle.
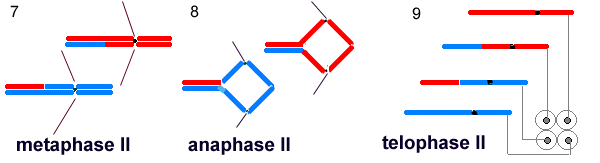
The chromosomes migrate to the equators of two new spindles for metaphase II (7). Next the chromatids are pulled apart in anaphase II to form four clusters of chromosomes in telophase II. The nuclear envelopes reform around four haploid nuclei that will give rise to the micro- or megagametophyte.
You can see "Virtual Meosis" online In a diploid organism each genetic locus is represented twice and may be occupied by identical copies of the gene (homozygous) or by copies (alleles) that differ in some way (heterozygous) When this happens we may see the expression of only one allele in the phenotype (dominance) - the other allele is recessive or sometimes we may see a blend of the two genes (incomplete dominance) When meiosis occurs in a heterozygous organism each of the gametes has an equal chance of getting one or other allele. In the simplest example of red (dominant) and white flowered peas cross pollination produces Ww (which is red) in the F1 generation. |
|
|
- |
|
|
|
eggs |
W |
w |
|
W |
WW |
Ww |
|
w |
Ww |
ww |
|
Thus we have three genotypes that will be represented by two phenotypes in the progeny in the following proportions: total WW.........25%........red Ww.........50%........red 75% red ww.........25%........white 25% white When dominance is incomplete there will be three phenotypes, corresponding to the genotypes. A similar cross in Antirrhinum produces 50% pink flowered plants in the F2 generation. This is the simplest type of segregation, resulting from a monohybrid cross. This situation allows us to illustrate the principle of a test cross to determine the genotype of a plant exhibiting the dominant phenotype (it could be WW or Ww). On crossing with the homozygous recessive parent the progeny could be: |
|
- |
|
|
|
eggs |
w |
w |
|
W |
red |
red |
|
? |
white |
white |
|
The occurrence of white progeny indicates that the unknown genotype must have included a recessive allele (w). If the progeny had been all red the parent would have been shown to be the homozygous dominant (WW). Mendel's first law (hereditable factors occur in pairs that can be separated in the process of sexual reproduction) is really a natural consequence of meiosis. It was not obvious at the time, because most people thought that characters were somehow blended in hybrids and could never be recovered in their original form. Mendel's second principle of independent assortment is also a consequence of meiosis in a dihybrid cross. When two loci such as R/r (round or wrinkled seed) and Y/y (yellow or green seed) are on different chromosomes, alleles will sort themselves out independently so that they can separate or go together at anaphase I or II of meiosis. A cross of RRYY with rryy produces RrYy (round and yellow) in the F1. This produces four types of gametes, male or female which can recombine in 16 ways to produce 9 genotypes and four phenotypes in the F2 generation. This is shown in a Punnet square: |
|
- |
|
|||
|
eggs |
RY |
Ry |
rY |
ry |
|
RY |
RRYY O |
RRYy O |
RrYY O |
RrYy O |
|
Ry |
RRYy O |
RRyy O |
RrYy O |
Rryy O |
|
rY |
RrYY O |
RrYy O |
rrYY |
rrYy |
|
ry |
RrYr O |
Rryy O |
rrYy |
rryy |
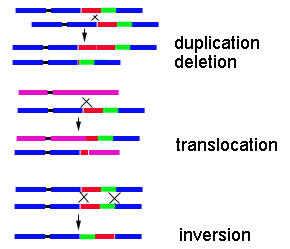
totals RRYY.........1 RRYy.........2 RrYY.........2 RrYy.........4 9 Round and Yellow Rryy.........1 Rryy.........2 3 Round and Green rrYY.........1 rrYy.........2 3 Wrinkled and Yellow rryy.........1 1 Wrinkled and Green Punnet squares can be constructed in an analogous fashion for three or more characters. Linkage and multigenic characters Although in principle any single genetic locus will undergo segregation, many plant characters are affected by more than one locus so that segregating populations show a continuous variation, rather than the discrete phenotypes which Mendel observed. (This gave people the idea of "blending"). Apparently Mendel never looked at hybrids involving pod shape and plant height which we now know to be on the same chromosome. In the analysis of the progeny of a dihybrid cross we noted that the characters were on different chromosomes. If they are linked i.e. on the same chromosome they will not show independent assortment; they will tend to stay together during meiosis. At the extreme they will always stay together; there will be a 1,2,1 ratio in the genotypes and only two phenotypes in 3:1 ratio. As the distance between loci increases the probability of cross-over or chiasma formation during meiosis increases, so that there will once again be four phenotypes. Opposite ends of a long chromosome may approach independent assortment so that 9:3:3:1 ratios of phenotypes are observed. The ratio of phenotypes is a measure of the distance between the genes on the chromosome and it is possible to build linkage maps of the chromosomes in this way. Mutation Mutations are caused by various kinds of high energy radiation (which can be natural), by chemical agents (which can be natural) or by mistakes in DNA replication or chromosome separation in mitosis or meiosis. Point mutations involve substitution of one or a few bases through replication errors. This may lead to amino acid changes in proteins which could be neutral or silent if they are not in a critical region of an enzyme.  This shows effects of different base changes in DNA on part of the amino acid sequence of a protein. Successive deletions are indicated by * Deletions could involve a single base or a large piece of a chromosome caused by radiation or errors in crossing over. Single base deletions disrupt the triplet code for amino acids (frameshift) so that the whole protein sequence is scrambled. Loss of one or more three-letter codons may not be so bad. Inversions of sections of DNA within a chromosome can occur by errors in ligation of broken ends during crossing over. This may have little effect on the growth of the progeny but there may be further problems during meiosis in the next generation. Translocations involve transfer of non-homologous pieces of DNA between chromosomes. They occur in the same way as inversions and the consequences are similar. Changes in chromosome number Occasionally in meiosis, chromosomes or chromatids may move to the wrong pole at anaphase I or II. One gamete may acquire an extra chromosome in its haploid set (monoploid) whereas another lacks a chromosome (aneuploid). Occasionally gametes may be formed without any reduction of chromosome number. Then a diploid pollen or egg fuses with its opposite number (which is haploid) to produce a zygote with three sets of chromosomes (triploid). Alternatively two diploid gametes may fuse so that the zygote has four sets of chromosomes (tetraploid). Plants are much more tolerant of such major chromosome alterations than most animals and such events seem to have been relatively common in plant evolution. |
|
|
This shows the consequences of errors in crossing-over after chromosome pairing during meiosis. |
|
|
Pleiotropy and Epistasis Phenotypic characters may require appropriate alleles of several genes in order to be expressed. Flower color is often determined by enzymes that hydroxylate anthocyanin molecules at various positions. However, if genes for one or more enzymes early in flavonoid metabolism are not functional, the pigment may not be formed at all. The flavonoid biosynthesis genes are epistatic to the hydroxylating enzyme genes.
Copyright © Michael Knee, |
|