
Populations | Population Genetics | Mutation Rate | Additional Sources of Variation
Natural Selection | Speciation | The Pace of Evolution | Evidence for Evolution | Links
Without variation (which arises from mutations of DNA molecules to produce new alleles) natural selection would have nothing on which to act. A population is a group of individuals living in the same geographical area and sharing a common gene pool. The gene pool is the sum of all genetic information carried by the members of a population.
All genetic variation in a population is generated by mutation. Mutation is any heritable change in DNA. Mutations can be changes of a single nucleotide base or may involve changes in chromosome number. Whether a mutation is good, neutral, or harmful depends on how it affects survival and reproductive success.
A population is a group of potentially interbreeding organisms of the same species occupying a certain area. Members of a population vary from one another. This variation is the raw material on which natural selection operates.
There are several types of mutations, both at the gene-level and the chromosome-level. Gene mutations provide new alleles, making these mutations the ultimate source of variation. A gene mutation is an alteration in the DNA nucleotide sequence, producing an alternate sequence, termed an allele. Mutations occur at random, and can be beneficial, neutral, or harmful. Some chromosomal mutations are changes in the number of chromosomes inherited, while others are alterations in arrangement of alleles on chromosomes due to inversions and translocations.
In sexually reproducing organisms, genetic recombination is the realloc ation of alleles and chromosomes. Recombination results from crossing-over during meiosis, the random segregation of chromosomes to gametes during meiotic division, and the random combination of gametes during fertilization. The entire genotype is subject to natural selection since new combinations of alleles may have improve the reproductive success of the organism. For polygenic traits, the most favorable combination may occur when the right alleles group by recombination.
Not only are variations created, they are also preserved and passed on from one generation to the next.The gene pool is the total of all the alleles in a population, in the context of gene frequencies. Neither dominance nor sexual reproduction will change allele frequencies.
This law states an equilibrium of allele frequencies in a gene pool (using a formula p2 + 2pq + q2) remains in effect in each succeeding generation of a sexually reproducing population if five conditions are met.
These conditions of the Hardy-Weinberg law are rarely met, so allele frequencies in the gene pool of a population do change from one generation to the next, resulting in evolution. We can now consider that any change of allele frequencies in a gene pool indicates that evolution has occurred. The Hardy-Weinberg law proposes those factors that violate the conditions listed cause evolution. A Hardy-Weinberg equilibrium provides a baseline by which to judge whether evolution has occurred. Hardy-Weinberg equilibrium is a constancy of gene pool frequencies that remains across generations, and might best be found among stable populations with no natural selection or where selection is stabilizing. Microevolution is the accumulation of small changes in a gene pool over a relatively short period.
Gene mutations result in new alleles, and are the source of variation within populations. Gene mutations are ultimately behind the other mechanisms that provide variation. Due to DNA replication and DNA repair mechanisms, mutation rates of individual genes are low, but since each organism has many genes, and a population has many individuals, new mutations arise in populations all the time. Thus, mutations are relatively common, and the mutation rate is an adequate source of new alleles. High levels of molecular variation are common in natural populations, although many mutations (usually recessive) are hidden.
The mutation rate varies greatly among species and even among genes of an individual. Mutations are caused by errors in DNA replication, chemicals, or radiation. Large scale effects of mutation result only when mutation is combined with other factors that reshuffle the gene pool.
Selection acts on individuals, not their individual genes. Sexual reproduction increases variation by reshuffling the genetic information from parents into new combinations in their offspring. Mutations produce new alleles.
Gene flow moves alleles among populations through interbreeding as well as by migration of breeding individuals. Gene flow increases variation within a population by introducing new alleles produced in another population. Continued gene flow tends to decrease the diversity among populations, causing gene pools to become similar. Reduction or restriction of gene flow between populations is essential for the development of new species.
The frequency of alleles can change from generation to generation as a result of chance alone in a small gene pool. This phenomenon is known as genetic drift.
Random mating involves individuals pairing by chance, not according to their genotypes or phenotypes. Nonrandom mating involves individuals inbreeding and assortative mating. Inbreeding is mating between relatives to a greater extent than by chance; inbreeding can occur if dispersal is so low that mates are likely to be related and does not change allele frequencies, but it does decrease the proportion of heterozygotes and increase the proportions of both homozygotes at all gene loci.
Assortative mating occurs when individuals tend to mate with those that have the same phenotype. Assortative mating divides a population into two phenotypic classes with reduced gene exchange.
Genetic drift is changes in allele frequencies of a gene pool due to chance or random events. This can occur in large or small populations. Genetic drift causes gene pools of two isolated populations to become dissimilar as some alleles are lost and other are fixed.
Genetic drift occurs when founders (or colonizers) establish a new population, or after a genetic bottleneck and resultant interbreeding. The founder effect is a case of genetic drift in which rare alleles, or combinations of alleles, occur in higher frequency in a population isolated from the general population. Founding individuals contain a fraction of the total genetic diversity of original gene pool. The alleles carried by founders is determined by chance alone. Consider the Pilgrim colonists in New England. By no means did they represent all the genetic variation of the human species or even genetic variations among Europeans.
When a population is started by one or a few individuals who randomly separate from a larger population, chance may dictate that allele frequencies in the new population may be very different from those of the original population. Many species on islands (such as the famous Darwin's finches on the Galápagos) display founder effects. The Galápagos Islands are volcanic islands off the coast of South America. They had fewer types of organisms than the South American mainland. The island species varied from the mainland species, and from island-to-island. Each island had a variation of tortoise that correlated with different vegetation and environmental conditions on that island.


Galapagos tortoises, note the difference in the height of the shell between the top and bottom images. Top image from Lycos, photo by Bill Everit; bottom image from http://home.capp.ch/marcel/Gal_Turt.htm.
Finches on the Galápagos Islands resembled a mainland finch but there were more types. Galápagos finch species varied by nesting site, beak size, and eating habits. One unusual finch used a twig or thorn to pry out insects, a job normally done by a woodpecker. The finches posed questions to Darwin: did they descend from one mainland ancestor, did islands allow isolated populations to evolve independently, and could present-day species have resulted from changes occurring in each isolated population.
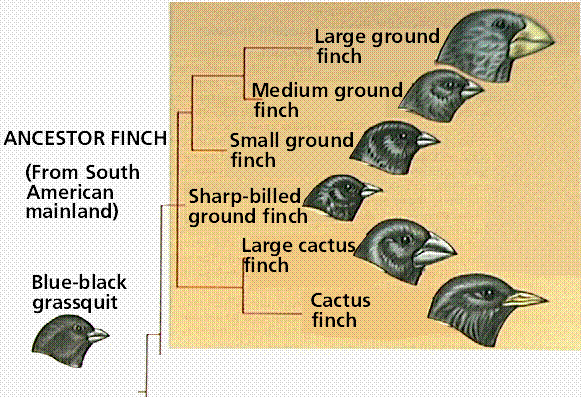
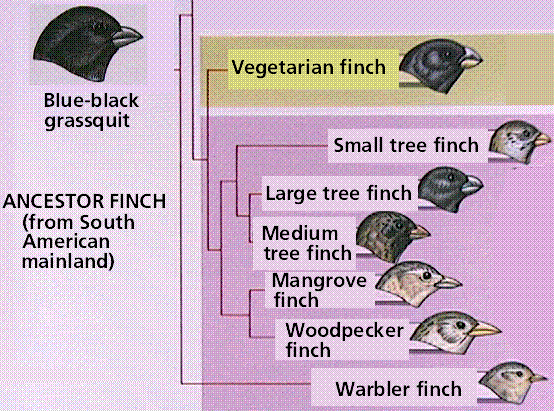
Divergence of the Galapagos finches from ancestral colonizers from the South American mainland. Images from W.H. Freeman and Sinauer Associates, used by permission.

Unidentified Galapagos finch.
Drastic short-term reductions of population size caused by natural disasters, disease, or predators may result in (by chance) the survivors representing only a small portion of the original gene pool. Even when the population increases to its original size, a portion of its original genetic diversity remains lost. This feature, termed a bottleneck, is a problem with many endangered species.
A bottleneck effect is genetic drift in which a severe reduction in population size results from natural disaster, predation, or habitat reduction. This results in a severe reduction of the total genetic diversity of the original gene pool. The cheetah bottleneck causes relative infertility because of the intense inbreeding. Similarly, the Hawaiian silversword has passed recently through its pwn bottleneck. Recent studies on humans suggest that there may have been one or more instances of severe genetic bottlenecks in our own prehistory. The bottleneck effect prevents most genotypes from participating in production of next generation.

Image of Hawaiian silverswords from http://mano.icsd.hawaii.gov/dlnr/images/silversword.jpg.
Migration into or out of a population can breakdown genetic differences between populations. Mutations developing in one population may be spread to other populations by migration. This serves, like mutation, to introduce new alleles into populations.
Not all members of a population necessarily have an equal chance of surviving and reproducing (due to competition for resources and mates). By virtue of small phenotypic variations, some individuals are better adapted to their environment than are others. The better adapted individuals are more "fit" and tend to survive and reproduce, passing on their adaptations to the next generation in greater frequency than those adaptations of the less "fit" members of the population.
Fitness is a measure of an individuals ability to survive and reproduce. Those with the highest fitness are more likely to survive and reproduce. Thus, they make a greater contribution to the gene pool, of the next generation than do those less "fit".
Natural selection is the process of differential survival and reproduction that inevitably leads to changes in allele frequencies over time as those individuals who are the most "fit" survive and leave more offspring. There are three patterns, or types, of natural selection.
Stabilizing selection favors the intermediate phenotype out of a range of phenotypes. The extremes in variation are selected against. Infants weighing significantly less or more than 7.5 pounds have higher rates of infant mortality. Selection works against both extremes.

Stabilizing selection, selection against the extremes in variation. Image from W.H. Freeman and Sinauer Associates, used by permission.
Living fossils, like the coelacanth, ginkgo, and horseshoe crab, are examples of organisms that are relatively unchanged from their distant ancestors.

Image of a coelacanth from http://www.dierentuin.net/pictures/coelacanth.jpg.

Exterior of a horseshoe crab, an example of a living fossil. From http://www.city.kasaoka.okayama.jp/horseshoe_crab/Kids_29.jpg.
Directional selection tends to favor phenotypes at one extreme of the range of variation. Insecticide resistance is an example. DDT was a widely used insecticide. After a few years of extensive use, DDT lost its effectiveness on insects. Resistance to DDT is a genetic trait that the presence of DDT in the environment made into a favored trait. Only those insects resistant to DDT survived, leading over time to populations largely resistant to DDT.
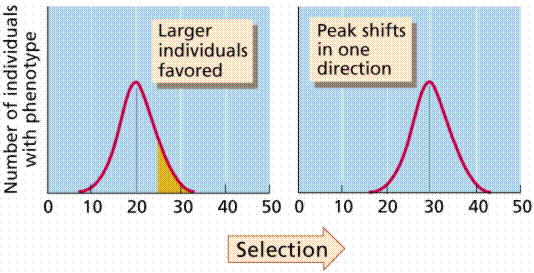
Image from W.H. Freeman and Sinauer Associates, used by permission.
Another example is the peppered moth (Biston betularia). Before the Industrial Revolution in the 18th and early 19th centuries, only light-colored moths were collected in light-colored woodlands in England. There was a rare, dark form. With the pollution caused by the buring of coal, the light-colored tree trunks became darker due to soot. The once rare dark-colored moths became more prevalent, while the once-common light-colored moths became increasingly rare. Reason: predation by birds. The color that had the greatest contrast with the background (tree trunk) was at a disadvantage. Cleanup of the forest during the 1950s caused the allele frequencies of light and dark moths to reverse to pre-Industrial Revolution levels, dark moths are now rare, light moths are now common.
The resistance of many bacterial species to antibiotics ia another example of directional selection. Over 200 speciews show some degree of antibiotic resistance, necessitating the development and more prudent use of a new generation of antibiotic medicines.
Disruptive selection favors individuals at both extremes of variation: selection is against the middle of the curve. This causes a discontinuity of the variations, causing two or more morphs or distinct phenotypes. The African swallowtail butterfly (Papilo dardanus) produces two distinct morphs, both of which resemble brightly colored but distasteful butterflies of other species. Each morph gains protection from predation although it is in fact quite edible.
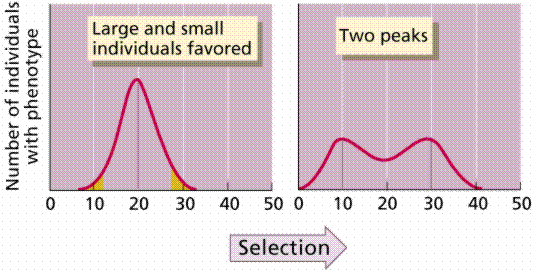
Image from W.H. Freeman and Sinauer Associates, used by permission.
As populations diverge, they form similar but related species. When are two populations new species? When populations no longer interbreed they are thought to be separate species. As natural selection adapts populations occupying different environments, they will diverge into races, subspecies, and finally separate species.
A species can be defined as one or more populations of interbreeding organisms that are reproductively isolated in nature from all other organisms. Genetic divergence results when adaptation, drift and mutation act on populations. Barriers to gene flow between populations isolate those populations, ultimately leading to the formation of new and separate species.
Populations begin to diverge when gene flow between them is restricted. Geographic isolation is often the first step in allopatric speciation. Other mechanisms may develop that further restrict reproduction between populations: these are the reproductive isolating mechanisms.
Sympatric speciation happens when members of a population develop some genetic difference that prevents them from reproducing with the parent type. This mechanism is best understood in plants, where failure to reduce chromosome number results in polyploid plants that reproduce successfully only with other polyploids. Reproduction with their parent population (the diploids) produces sterile offspring.
A reproductive isolating mechanism is a structural, functional, or behavioral characteristic that prevents successful reproduction from occurring. These mechanisms divide into premating and postmating types.
Premating isolating mechanisms are anatomical or behavioral differences between two species that prevent the possibility of mating. Habitat isolation occurs when two species occupy different habitats, even within the same geographic range, so that they are less likely to meet and to attempt to reproduce. Temporal isolation occurs when two species live in the same location, but each reproduces at a different time of the year, preventing a successful mating. Behavioral isolation occurs when there are differences in mating behavior between two species. Mechanical isolation is the result of differences between two species in reproductive structures or other body parts, so that mating is prevented.
Postmating isolating mechanisms are the result of developmental or physiological differences between the members of two species after mating. Gamete isolation is the physical or chemical incompatibility of gametes of two different species. If the gametes lack receptors to facilitate fusion, they cannot form a zygote. An egg may have receptors only for the sperm of its own species. Zygote mortality is a mechanism that works when hybrids (offspring of parents of two different species) do not live to reproduce. Hybrid sterility occurs when the hybrid offspring are sterile (e.g., mules).
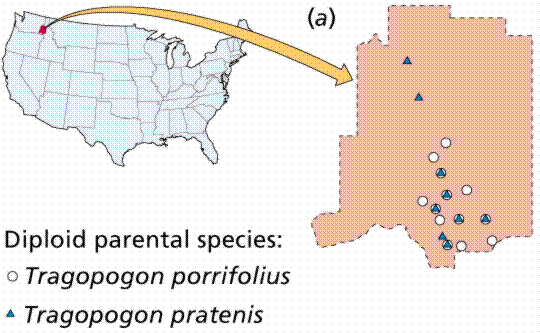
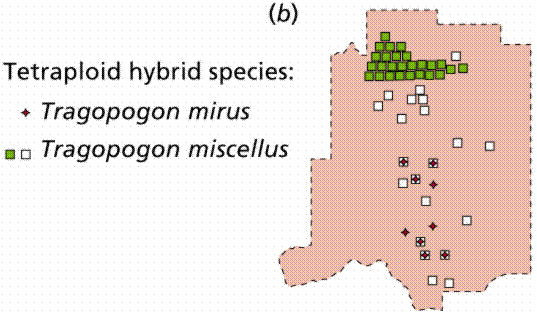
Image from W.H. Freeman and Sinauer Associates, used by permission.
Polyploidy and hybridization are important speciation mechanisms in plants. Whereas animals tend to be unisexual, plants often have both sexes functional in the same individual. Consequently, plants can (if they lack a self-incompatability mechanism) reproduce with themselves (both sexually and asexually), establishing a reproductively isolated species very rapidly.
The pace of evolution is often slow, so slow that all of the stages in species formation cannot be observed. The traditional, or Darwinian, view of evolution was that it was a very slow process, resulting from the gradual accumulation of small differences. Recently, several alternative views on the pace and events in species formation have been proposed. These are collectively called quantum speciation.
According to another model based on the fossil record, speciation occurs rapidly over a short time, followed by a long period of little or no change. "Short" means thousands or hundreds of thousands of years. This differs greatly from Darwin's original view of slow and gradual change continuing over very long periods of time.
Evolution, which started out as a hypothesis, is now supported by evidence from many fields of science.
The fossil record is the history of life recorded by remains from the past. Fossils include skeletons, shells, seeds, insects trapped in amber, dung, DNA and other chemicals, imprints of leaves, and tracks of organisms that lived in the distant past. Most fossils are at least 10,000 years old.
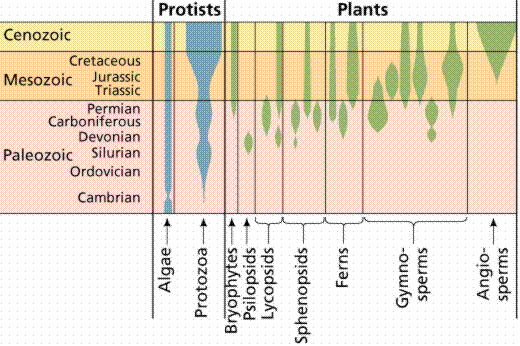
The fossil records of some protist and plant groups. The width of the shaded space is an indicator of the number of species. Image from W.H. Freeman and Sinauer Associates, used by permission.
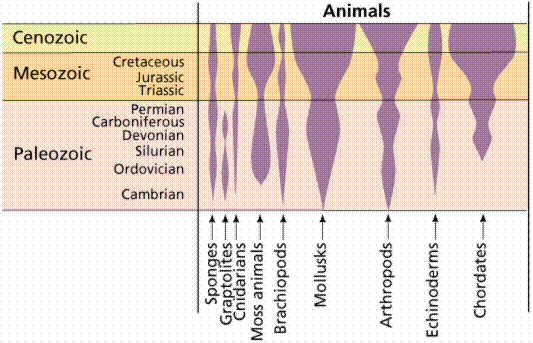
First appearances and relative diversity (width of shaded area) for major groups of animals. Image from W.H. Freeman and Sinauer Associates, used by permission.
The fossil record traces history of life and allows us to study history of particular organisms. Fossil evidence supports the common descent hypothesis; fossils can be linked over time because they reveal a similarity in form, despite observed changes. Transitional forms reveal links between groups: Archaeopteryx is between reptiles and birds; Eustheopteron is an amphibious fish; Seymouria is a reptile-like amphibian; therapsids were mammal-like reptiles.

These images of Archaeopteryx, once considered the first bird, are from http://www.ucmp.berkeley.edu/diapsids/birds/. The fossil is from the Solenhoefen Limestone (Jurassic) of Germany.
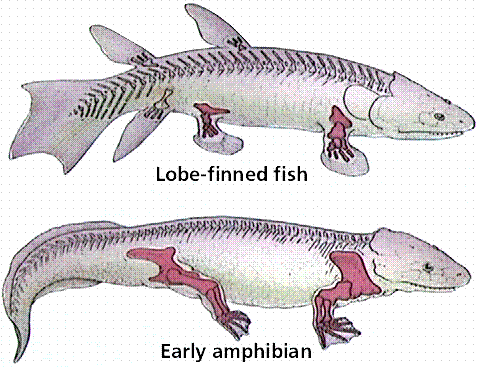
Comparison of the skeletons of a crossopterygian lobe-finned fish and an early amphibian. Image from W.H. Freeman and Sinauer Associates, used by permission.
The fossil record allows us to trace the history of the modern-day horse Equus. The earliest fossils in this lineage is Hyracotherium , which was the size of a dog, with cusped low-crowned molars, four toes on each front foot, three on each hind foot--all adaptations for forest living. When forests were replaced by grasslands, the intermediates were selected for durable grinding teeth, speed, etc. with an increase in size and decrease in toes. Living organisms resemble most recent fossils in the line of descent; underlying similarities allow us to trace a line of descent over time.
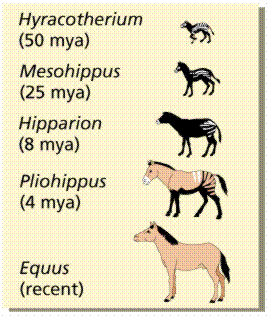
Stages in the evolution of the horse. Image from W.H. Freeman and Sinauer Associates, used by permission.
Biogeography is the study of the distribution of plants and animals throughout the world. Distribution of organisms is explained by related forms evolving in one locale and spreading to other accessible areas. Darwin observed South America had no rabbits; he concluded rabbits originated elsewhere. Biogeography explains why many finch species are on the Galápagos Islands but not mainland.
Physical factors, such as the location of continents, determine where a population can spread. Cacti are restricted to North American deserts and euphorbia grow in African deserts. Marsupials arose when South America, Antarctica, and Australia were all joined; Australia separated before placental mammals arose, so only marsupials diversified in Australia.
Organisms have anatomical similarities when they are closely related because of common descent, as substantiated by comparative anatomy. Homologous structures in different organisms are inherited from a common ancestor. Vertebrate forelimbs contain the same sets of bones organized in similar ways, despite their dissimilar functions.
Vestigial structures are remains of a structure that was functional in some ancestor but is no longer functional in the organism in question. Most birds have well-developed wings, although some birds have reduced wings and do not fly. Humans have a tail bone (the coccyx) but no tail. The presence of vestigial structures is explained by the common descent hypothesis.
Embryological development reveals a unity of plan. During development, all vertebrates have a notochord and paired pharyngeal pouches. In fishes and amphibian larvae, the pouches become gills. In humans, first pair of pouches becomes a cavity of middle ear and auditory tube; second pair becomes tonsils, while third and fourth pairs become thymus and parathyroid glands. This makes sense only if fish are ancestral to other vertebrate groups.
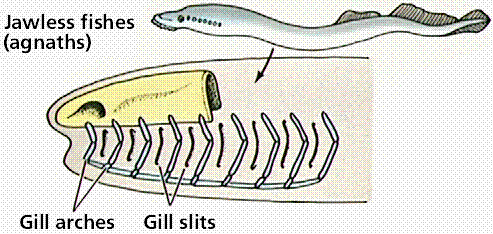
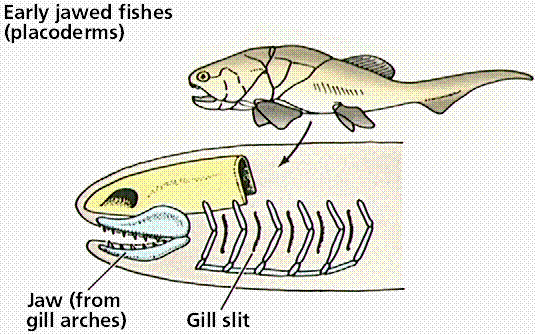

Steps in the evolution of jaws by modification of gill arches. Images from W.H. Freeman and Sinauer Associates, used by permission.
Almost all living organisms use the same basic biochemical molecules, including DNA, ATP, and many identical or nearly identical enzymes. Organisms utilize the same DNA triplet base code and the same 20 amino acids in their proteins. Many organisms share same introns and types of repeats, which is remarkable since we know of no obvious functional reason why these components need to be so similar. These similarities can be explained by descent from a common ancestor. This is substantiated by analysis of degree of similarity in amino acids for cytochrome c among organisms.
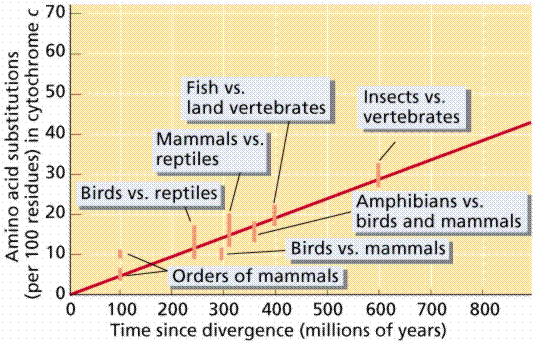
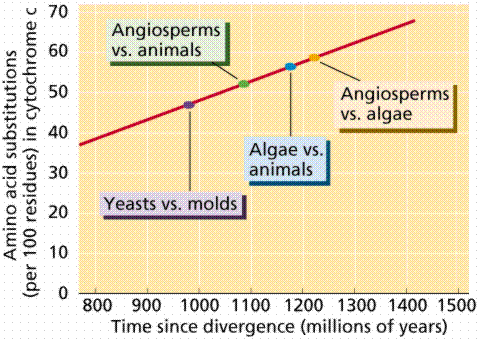
Interpretation of results of biochemical study of cytochrome c among the eukaryotes. Images from W.H. Freeman and Sinauer Associates, used by permission.
Email: mj.farabee@emcmail.maricopa.edu![]()
Last modified: 2000/01/07:13:26:00
The URL of this page is: gened.emc.maricopa.edu/bio/BIO181/BIOBK/BioBookEVOLII.html
{kind=link}
{kind=link}
{kind=link}