

Nuphar lutea - Gelbe Teichrose (Aufn. K. HANDKE)
Bevor wir uns ausführlich mit dem Bau von Blüten befassen, sei vorausgeschickt, daß es sich bei ihnen um Strukturen handelt, die der sexuellen Fortpflanzung dienen; dabei entstehen als Endprodukt Samen. Die Blütenpflanzen werden daher auch Samenpflanzen (Spermatophyta) genannt. Sie umfassen zwei große Gruppen, die Gymnospermen (Nacktsamer) und die Angiospermen (Bedecktsamer). Die Gymnospermen sind die stammesgeschichtlich ältere und primitivere Gruppe. Im streng systematischen Sinne darf man sie nicht als Klasse bezeichnen, denn ihr gehören zwei echte Klassen, die Koniferen (Nadelbäume) und die Cycadeen an, welche nicht auf gleiche Vorfahren zurückzuführen sind. Die Angiospermen, die man vielfach als die eigentlichen Blütenpflanzen (Blütenpflanzen im engeren Sinne) ansieht, gehören nur einer Klasse an, sie repräsentieren die überwältigende Mehrheit der Arten; und das, was wir im folgenden besprechen werden, bezieht sich in vornehmlich auf ihre Blüten.
Es darf wohl vorausgesetzt werden, daß jeder aufgrund eigener Erfahrung einen Eindruck von der Mannigfaltigkeit der Blütenformen, ihrer Farben und Düfte gewonnen hat. Es ist sicher auch bekannt, daß diese Merkmale im Verlauf der Evolution in Anpassung an bestimmte Bestäuber (Insekten: Bienen, Hummeln. Schmetterlinge, Fliegen u.a., Vögel: Kolibris, Honigvögel u.a.) entstanden sind. Windbestäubte Pflanzen, zu denen in erster Linie die Gymnospermen. aber auch viele Angiospermen gehören, haben unauffällige Blüten.
In vielen Schulbüchern werden die Blüte der Anemone (Anemone nemorosa) und/oder der Tulpe (Tulipa gesneriana) als typisches Beispiel für sie vorgestellt. Die Wahl ist nicht schlecht, denn an deren Blüten kann der Grundbauplan besser demonstriert werden als an vielen anderen.
Blüten sind Kurztriebe mit begrenztem Wachstum. An einer meist stark gestauchten Blütenachse sind in schraubiger oder wirteliger Anordnung in mehreren Kreisen (Wirteln) übereinander Blattorgane angeordnet, die sich in ihrem Aussehen und ihrer Funktion deutlich von den übrigen (Laub-) Blättern abheben.
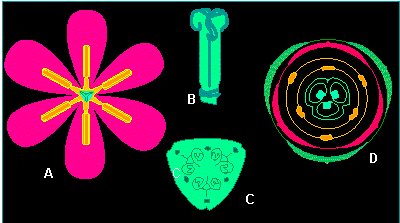
Tulipa gesneriana (Tulpe). A. Blüte in Aufsicht, B und C. Pistill (Stempel) Gesamtansicht (B), Querschnitt im Bereich des Ovars (C); D. generalisiertes Blütendiagramm. Blütendiagramme wurden im vergangenen Jahrhundert von dem Kieler Botaniker A. EICHLER (1875, 1878) für die Mehrzahl der einheimischen und fremdländischen Pflanzengruppen erstellt. Sie symbolisieren die Position, die Zahl und die Symmetrieverhältnisse der einzelnen Komponenten einer Blüte in den jeweiligen Blütenblattkreisen (Nach W. TROLL, 1975)
Von unten nach oben (bzw. von außen nach innen) fortschreitend, unterscheidet man dabei zwischen dem meist unscheinbar gefärbten Kelch (Calyx, aus Sepalen bestehend), der meist auffällig gefärbten Blumenkrone (Corolla, aus Petalen), den Staubblättern (Stamina) und schließlich den Fruchtblättern (Karpellen). Kelch und Blumenkrone zusammen bilden die Blütenhülle, das Perianth. Wenn sich Kelch und Blumenkrone jedoch nicht voneinander unterscheiden lassen oder wenn der Kelch fehlt, liegt eine einfache Blütenhülle (Perigon) vor. Fehlt die Blütenhülle ganz, spricht man von nackter Blüte. Solche Blüten sind für windbestäubte Arten typisch.
Die Gesamtheit der Staubblätter (männliche Fortpflanzungsorgane) bezeichnet man als Androeceum, die der Fruchtblätter (weibliche Fortpflanzungsorgane) als Gynoeceum.
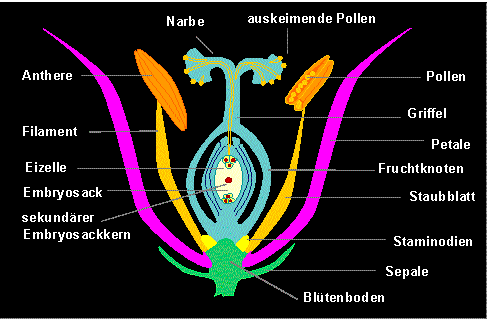
Die Blütenachse tritt bei den meisten Angiospermen kaum in Erscheinung, sie ist gewöhnlich verkürzt und zu einem sog. Blütenboden verbreitert; gelegentlich ist er leicht gewölbt oder scheibenförmig ausgebildet (Diskus).
Bei den Gymnospermen hingegen sowie bei manchen der primitiven Angiospermen (z.B. Magnolia) ist sie in Form eines ansehnlichen Achsenzapfens ausgebildet .
Ursprünglich besaßen die Blüten der Angiospermen nur eine einfache Blütenhülle, die wahrscheinlich auf Hochblätter zurückzuführen ist, und die in sehr einfacher Form schon bei den Gymnospermenblüten vorkommt. Die Mehrzahl der Angiospermenblüten besitzt jedoch, wie bereits geschildert, eine doppelte Blütenhülle mit einer klaren Aufgabentrennung zwischen Kelch und Krone. Während die Krone in der Regel die Schaufunktion (d.h. die Anlockung der Bestäuber) übernimmt, dient der Kelch primär dem Schutz der heranwachsenden übrigen Blütenorgane. Er verliert seine Bedeutung mit der Blütenentfaltung (Anthese); bei manchen Arten, z.B. dem Mohn, werden die Kelchblätter dann abgestoßen, bei den meisten anderen stellt der Kelch sein Wachstum nach der Entfaltung der Krone ein und vertrocknet. Nur selten werden die Kelchblätter nach der erfolgreichen Bestäubung der Blüte reaktiviert; ein erneutes Wachstum setzt ein, wodurch sich eine Hülle um die reifende Frucht bildet.

radiärsymmetrisch |
 Ursprünglich sind alle Blütenorgane in einer engen Schraube
angeordnet gewesen (z.B. Anemone). Diese Blüten sind strenggenommen asymmetrisch, werden aber in den Bestimmungsbüchern zu den
radiärsymmetrischen
(aktinomorphen oder strahligen) Blüten gestellt. Gekennzeichnet sind sie durch die Anordnung ihrer Blütenorgane auf Wirteln (in Blütenkreisen),
wobei alle Organe innerhalb eines Wirtels gleichgestaltet sind und jeder Wirtel die gleiche Zahl von Organen aufweist (die Zahlen drei, vier und fünf sind sehr häufig).
Ursprünglich sind alle Blütenorgane in einer engen Schraube
angeordnet gewesen (z.B. Anemone). Diese Blüten sind strenggenommen asymmetrisch, werden aber in den Bestimmungsbüchern zu den
radiärsymmetrischen
(aktinomorphen oder strahligen) Blüten gestellt. Gekennzeichnet sind sie durch die Anordnung ihrer Blütenorgane auf Wirteln (in Blütenkreisen),
wobei alle Organe innerhalb eines Wirtels gleichgestaltet sind und jeder Wirtel die gleiche Zahl von Organen aufweist (die Zahlen drei, vier und fünf sind sehr häufig).
Die aufeinanderfolgenden Wirtel stehen meist auf "Lücke" (Regel von der Alternanz). Durch solche Blüten können zwei oder mehr Symmetrieebenen gelegt werden (Blüten mit nur zwei Symmetrieebenen werden manchmal auch als bilaterale Blüten unterschieden). Selbst wenn das Gynoeceum eine abweichende Zahl von Organen aufweisen sollte, werden die Blüten zu den radiärsymmetrischen gerechnet.
 Weit verbreitet sind Blüten, durch die sich aufgrund unterschiedlicher Ausgestaltung der Blütenorgane innerhalb eines Wirtels oder aufgrund
unvollständiger Organwirtel (z.B. Wirtel mit einer geringeren Organzahl, besonders häufig im Androeceum) nur noch eine Symmetrieebene legen läßt. Diese als besonders abgeleitet geltenden Blüten nennt man als zygomorph (monosymmetrisch oder
dorsiventral). Beispiele hierfür findet man beim Stiefmütterchen (Viola) oder den Lippenblüten der Lippenblütler (Lamiaceae).
Weit verbreitet sind Blüten, durch die sich aufgrund unterschiedlicher Ausgestaltung der Blütenorgane innerhalb eines Wirtels oder aufgrund
unvollständiger Organwirtel (z.B. Wirtel mit einer geringeren Organzahl, besonders häufig im Androeceum) nur noch eine Symmetrieebene legen läßt. Diese als besonders abgeleitet geltenden Blüten nennt man als zygomorph (monosymmetrisch oder
dorsiventral). Beispiele hierfür findet man beim Stiefmütterchen (Viola) oder den Lippenblüten der Lippenblütler (Lamiaceae).

Bei den meisten Blütenpflanzen sind die Kronblätter getrennt (freikronblättrige Blüten oder Dialypetalae). Bei den verwachsenkronblättrigen (Sympetalae) sind sie, die Petalen, untereinander zu einer Kronröhre vereint. Aus der Zahl der freien Zipfel kann die Zahl der verwachsenen Petalen erschlossen werden.
Die Symmetrieverhältnisse und der Bau der Blüten läßt sich am anschaulichsten durch Blütendiagramme wiedergeben. Sie repräsentieren schematisierte Grundrisse, in denen die einzelnen Organe (Zahl und Position) wiedergegeben sind. Beim Vergleich der Diagramme zeigt sich, daß bei den Dikotyledonen die Grundzahl 5 für die einzelnen Wirtel typisch ist, während bei den Monokotyledonen die Zahl drei vorherrschend ist (Trimerie). Radiärsymmetrische Blüten werden aktinomorph genannt, Blüten mit einer Symmetrieebene zygomorph.

Die Kronblätter (Petalen) lassen sich entwicklungsgeschichtlich vielfach auf umgebildete Staubblätter zurückführen. Diese Deutung ist durch das wiederholt nachgewiesene Auftreten von Zwischenformen der beiden Organkategorien begründbar; ein klassische Beispiel ist die Weiße Seerose (Nymphaea alba, übrigens eine recht primitive Angiosperrne), doch findet man solche Metamorphosen auch bei vielen anderen Arten. Züchter nutzen diese Eigenschaft zur Selektion "gefüllter" Blüten, so besitzt die wildlebende Heckenrose (Rosa canina) nur 5 Kronblätter und viele Staubblätter, während die meisten der heutigen Kultursorten sich durch "volle" Blüten mit zahlreichen Kronblättern und einer verminderten Zahl (fertiler) Staubblätter auszeichnen. Windbestäubte Blüten zeichnen sich vielfach durch das Fehlen von Kronblättern aus.
Ein normal ausgebildetes Staubblatt setzt sich aus einem fadenförmigen, sterilen Staubfaden (Filament) und einem fertilen Staubbeutel (Anthere) zusammen. Die Anthere wiederum besteht aus (meist) zwei, durch ein Konnektiv verbundene Theken. Jede Theka enthält zwei Pollensäcke, in denen die Pollenbildung erfolgt.
Unfruchtbare Staubblätter nennt man Staminodien. Eine ihrer Varianten sind die nektarabsondernden Honigblätter, die oft zwischen dem Perianth und den Staubblättern liegen. Sie können die Größe und Färbung von Kronblättern annehmen und den Schauapparat der Blüte bilden (so z.B. beim Hahnenfuß, Ranunculus, Ranunculaceae).
Den zentralen Teil einer typischen Angiospermen-Blüte bildet das aus einem oder mehreren Fruchtblättern (Karpellen) bestehende Gynoeceum. Die Fruchtblätter sind - im Gegensatz zu denen der Gymnospermen - geschlossen (s.u.); ihr Blattcharakter ist daher nur noch sehr undeutlich. Das Gynoeceum umfaßt einen oder mehrere Stempel.
Ein typischer Stempel besteht (von unten nach oben) aus dem Fruchtknoten (Samenanlagen enthaltend), dem Griffel und der Narbe (Stigma), dem Aufnahmeorgan (Empfängnisorgan) für Pollenkörner. Narben können sehr variabel strukturiert sein. Oft sind sie knopfförmig und mit Papillen versehen, nicht selten sind sie verzweigt, wobei einzelne Narbenäste erkennbar sind. Man unterscheidet zwischen trockenen und feuchten Narben. Die Oberfläche der trockenen ist meist mit zu haarförmigen Papillen ausgezogenen Zellen besetzt, die der feuchten ist mit einer klebrigen Schicht überzogen.
Der Griffel dient dazu, die Narbe in eine für die Bestäubung günstige Position zu bringen.
Bei einem Querschnitt durch den Fruchtknoten kann man den Blattcharakter der Karpelle oft recht gut erkennen. Die beiden Ränder eines Fruchtblattes erscheinen eingerollt und miteinander verwachsen, so daß eine röhrenförmige Struktur entsteht, an deren Grunde innerhalb des so gebildeten Hohlraums (geschützt durch das eingerollte Fruchtblatt: "bedeckt") die Samenanlagen liegen. Bei den Gymnospermen fehlt dieser Schutz; die Samenanlagen liegen dort frei auf den Fruchtblättern.
Bei einem Angiospermenfruchtblatt wird die durch Verwachsung der Blattränder gebildete Naht als Bauchnaht bezeichnet; die Mittelrippe der Karpelle ist (etwas irreführend bezeichnet) die Rückennaht.
Üblicherweise umfaßt das Gynoeceum der Angiospermenblüte mehrere Karpelle. Eine bemerkenswerte Ausnahme bilden die Leguminosen (Hülsenfrüchtler), bei denen die Blüte nur ein einziges Karpell enthält. Die Fruchtblätter eines Gynoeceums können entweder jedes für sich einen Stempel bilden (Chorikarpie oder Apokarpie) oder mehrere Fruchtblätter können zu einem Stempel verwachsen sein (Coenokarpie); der Vorteil liegt darin, daß nach einem einzigen erfolgreichen Bestäubungsvorgang die Pollenschläuche auf alle Karpelle verteilt werden können.
Die Art der Verwachsung der Karpelle und damit die Lage der Samenanlagen innerhalb des Fruchtknotens, die sog. Placentation, wurden von A. ENGLER und E. PRANTL Ende des vergangenen Jahrhunderts als wichtiges taxonomisches Merkmal zur Untergliederung der Angiospermen erkannt. So wurden einige Ordnungen (Centrospermae, Parietales) nach der Art der Fruchtknotenarchitektur benannt.
Ein aus freien Karpellen bestehender Fruchtknoten ist apokarp (apokarpes oder chorikarpes Gynoeceum) und stellt die ursprünglichste Form des Angiospermengynoeceums dar; bei Ranunculaceen ist diese Form oft zu finden. Abgeleitet sind hiervon die Fruchtknoten mit verwachsenen Fruchtblättern.
Bei einem synkarpen Fruchtknoten sind die Karpelle seitlich miteinander fusioniert. Er hat eine der Zahl der Karpelle entsprechende Zahl von Kammern (sog. Fächern), die durch Scheidewände getrennt sind, was sich besonders klar an Querschnitten durch den Fruchtknoten einer Tulpe demonstrieren läßt. Die Samenanlagen sind im inneren Winkel der Fächer inseriert (zentralwinkelständige oder axile Placentation). Zusätzlich kann jedes Fach durch eine radial verlaufende, sog. falsche Scheidewand untergliedert sein.
Bei einem parakarpen Fruchtknoten hingegen sind die benachbarten Karpelle mit ihren Rändern verwachsen und umschließen daher einen einheitlichen Hohlraum. Die Samenanlagen sitzen dort an den Verwachsungsnähten der Karpelle auf der Innenseite der Fruchtknotenwand (wandständige oder parietale Placentation). Auch hier können nachträglich falsche Scheidewände eingezogen sein, was bei oberflächlicher Betrachtung zu dem Trugschluß führen könnte, man habe es mit einem synkarpen Fruchtknoten zu tun. Die unverändert parietale Placentation der Samenanlagen ermöglicht jedoch eine eindeutige Zuordnung.
Einen einheitlichen Hohlraum - bedingt durch die Auflösung der Seitenränder - findet man bei den lysikarpen Fruchtknoten, hier sind die Samenanlagen jedoch auf einem Achsenzapfen im Zentrum des Fruchtknotens inseriert (freie Zentralplacenta, frei zentrale Placentation).
Die Position des Fruchtknotens in Relation zu den übrigen Blütenorganen ist ein wichtiges taxonomisches Merkmal. Zu unterscheiden ist zwischen oberständigem, mittelständigem und unterständigem Fruchtknoten (hypogyn, perigyn und epigyn).
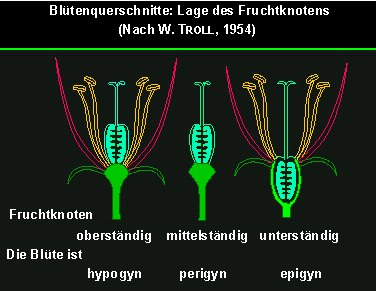
Von links nach rechts: oberständiger, Mittelständiger und unterständiger Fruchtknoten. Die Blüten sind demnach hypogyn, perigyn und epigyn.
Blüten, die sowohl ein Androeceum als auch ein Gynoeceum enthalten, nennt man zwittrig oder hermaphrodit. Enthalten sie nur ein Androeceum, haben wir männliche Blüten vor uns; enthalten sie nur ein Gynoeceum, haben wir es mit weiblichen Blüten zu tun. Befinden sich männliche und weibliche Blüten auf einer Pflanze, ist sie einhäusig oder monözisch, sind sie auf zwei Individuen verteilt, sind die Pflanzen zweihäusig oder diözisch ( Monözie und Diözie).
Im Extremfall kann eine Blüte auf ein einzelnes Staubblatt oder ein einzelnes Fruchtblatt reduziert sein. Einen besonders interessanten Fall findet man bei der Wolfsmilch (Euphorbia) und einigen nahen Verwandten. Hier sind solche extrem reduzierten Blüten zu einer funktionell zwittrigen "Scheinblüte" - Cyathium genannt - zusammengefaßt. Durch Integration eines Satzes von Hochblättern, die große, meist gelb gefärbte Nektarien tragen, und u. U. weiteren auffällig gefärbten Hochblättern, wird der Eindruck einer normalen zwittrigen Angiospermenblüte perfektioniert, z.B. beim schon erwähnten Weihnachtsstern (Poinsettia pulcherrima).
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de