
Die 3. MENDELsche Regel postuliert, daß Anlagen für die verschiedenen Merkmale unabhängig voneinander vererbt werden. Die Aussage trifft für alle sieben von ihm analysierten Merkmale zu. Wie er aber in seinem Briefwechsel mit C. v. NÄGELI schon andeutete, wird die Regel gelegentlich durchbrochen. Einen Beleg dafür erbrachten im Jahre 1905 W. BATESON, E. R. SAUNDERS und R. C. PUNNETT durch Arbeiten an der Spanischen Wicke (Lathyrus odoratus). Untersucht wurde die Vererbung von Blütenfarbe und Pollenstruktur. 381 F2-Pflanzen wurden ausgewertet. Die entsprechenden Merkmale verteilten sich wie folgt:
284 (=74,6%) Blütenfarbe: purpur, Pollen: lang
21 (=5,5%) Blütenfarbe: purpur, Pollen: rund
21 (=5,5%) Blütenfarbe: rot, Pollen: lang
55 (= 14%) Blütenfarbe: rot, Pollen: rund
In einem Wiederholungsexperiment fand R. C. PUNNETT (1917) unter 6952 F2-Pflanzen:
4831 (=69,5%) Blütenfarbe: purpur, Pollen: lang
390 (=5,6%) Blütenfarbe: purpur, Pollen: rund
393 (=5,6%) Blütenfarbe: rot, Pollen: lang
1338 (= 19,3%) Blütenfarbe: rot, Pollen: rund
Das bedeutet also, daß die Merkmalsanlagen in der Regel gemeinsam vererbt werden. Sie werden bei der Gametenbildung normalerweise also nicht voneinander getrennt, so daß die Kombinationen Blütenfarbe purpur, Pollen lang: Blütenfarbe rot, Pollen rund, im ungefähren Verhältnis 3:1 auftreten. Bemerkenswert sind die Ausnahmen, denn gelegentlich wird die Kopplung durchbrochen, und es treten in geringen Prozentzahlen die reziproken Kombinationen (Blütenfarbe purpur, Pollen rund) und, in gleicher Menge, Blütenfarbe rot, Pollen lang, auf. Die Kreuzungsexperimente geben keine schlüssige Erklärung für diese Befunde. Doch unter Einbeziehung der Chromosomentheorie erhält man eine durchaus plausible Antwort.
Spätere Untersuchungen von T. H. MORGAN und seinen Mitarbeitern an der Taufliege Drosophila melanogaster ergaben, daß die Zahl der Kopplungsgruppen der Anzahl der Chromosomen im haploiden Satz entspricht und daß die gelegentlichen Ausnahmen auf einem Chromosomenstückaustausch beruhen, der während der Paarung homologer Chromosomen in der Meiose erfolgt. Die Daten aus Kreuzungsexperimenten wurden einerseits zu einer wichtigen Stütze der Chromosomentheorie, andererseits eröffnete die Auswertung der Austauschhäufigkeiten einen Weg, Gene auf Chromosomen zu lokalisieren.
Bereits wenige Jahre nach der Wiederentdeckung der MENDELschen Regeln beschrieben C. CORRENS (1909) und E. BAUR (1909, 1910) Kreuzungsexperimente, auf die die MENDELschen Regeln nicht anwendbar waren. So wird ein bestimmter Typ der Blattscheckigkeit (grün-weiß-Panaschierung von Mirabilis jalapa und Antirrhinum majus sowie einigen anderen Arten nach dem folgenden Schema vererbt:
(1) (weiblich) grün x (männlich) grün-weiß gescheckt
Fl ausschließlich grüne Pflanzen
(2) (weiblich) grün-weiß gescheckt x (männlich) grün
F1: ausschließlich grün-weiß gescheckte Pflanzen.
Die Reziprozitätsregel, also praktisch die 1. MENDELsche Regel ist damit verletzt. In der F2 kommt es zu keinerlei zahlenmäßig erfaßbaren Aufspaltung. Es entfällt somit auch die Gültigkeit der 2. MENDELschen Regel.
Für die Ausprägung des Phänotyps der F1 ist nur der mütterliche Genotyp ausschlaggebend. Im Gegensatz zum Kern des Pollens, ist der Kern der Eizelle von einem voluminösen Plasma umgeben; es lag daher der Gedanke nahe, daß dieses Plasma Erbträger enthält, die sich bei der beobachteten Verteilung von weiß-grün- und grün-Phänotypen auswirken. Folglich nannte man diese Erscheinung plasmatische oder mütterliche, später auch extrachromosomale Vererbung.
Bei Pelargonium zonale stellte E. BAUR eine Variante dieses Vererbungsmodus fest: Es gibt von dieser Art, außer Pflanzen mit rein grünen Blättern, weißgeränderte und gelbgeränderte (Weißrand- und Gelbrand-Pelargonien). Die Vererbung der Gelbrandform erfolgt nach dem gleichen Muster wie die der Weißrandform. In der F1 können bereits die Kotyledonen ausgewertet werden. Dabei treten, neben grünen und gescheckten, auch einige rein weiße Formen auf, die sich wegen des Fehlens von Chlorophyll natürlich nicht weiterentwickeln können. Die Ergebnisse der Kreuzungsexperimente:
(1) (weiblich) grün x (männlich) grün-weiß gescheckt
F1: in drei voneinander unabhängigen Ansätzen wurden 159
Exemplare ausgewertet, davon waren: 138 grün, 17 gescheckt, 4 weiß
(2) (weiblich) grün-weiß gescheckt x (männlich) grün
Fl: 65 Exemplare wurden ausgezählt, davon waren: 46 grün, 19 gescheckt und 0 weiß
Dieses Beispiel ähnelt zwar den Beobachtungen an Mirabilis jalapa und Antirrhinum majus, macht aber zugleich deutlich, daß auch das (wenige) Plasma des Pollens einen Einfluß auf die Merkmalsausprägung haben kann.
Was sind plasmatische Faktoren? Welche Bedingungen müssen Erbträger erfüllen? Erbträger müssen ganz allgemein zwei Kriterien erfüllen:
| sie müssen die sie enthaltende Information weitergeben und | |
| sie müssen in der Lage sein, sich selbst zu verdoppeln (replizieren). Verlorengegangene Erbträger sind nicht ersetzbar. |
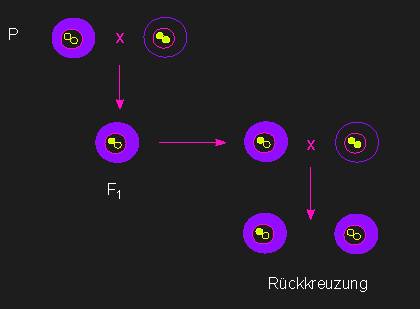
Nachweis plasmatischer Vererbung. Der Phänotyp ist unabhängig vom Kerngenom.
Wie schon an anderer Stelle vermerkt, erfüllen außer den Chromosomen auch die Plastiden diese Bedingungen. Ihre morphologische Kontinuität wurde in den Jahren 1882-1885 von F. SCHMITZ, A. F. W. SCHIMPER und A. MEYER erwiesen und hierauf aufbauend deutete E. BAUR (1909) seine Ergebnisse wie folgt:
"Die befruchtete Eizelle, die entstanden ist durch Vereinigung einer "grünen" mit einer "weißen" Sexualzelle enthält demnach zweierlei Chromatophoren (Anm.: Plastiden), grüne und weiße. Bei den Zellteilungen der zum Embryo auswachsenden Eizelle verteilen sich die Chromatophoren ganz nach Zufallsgesetzen auf die Tochterzellen. Erhält eine Tochterzelle nur weiße Chromatophoren, so wird diese Zelle weiterhin nur weiße Zelldeszendenzen (Anm.: Nachkommen in der betreffenden Zellinie) haben und ein weißes Mosaikstück aus sich hervorgehen lassen. Erhält eine Tochterzelle nur grüne Chromatophoren, so entsteht daraus ein konstant grüner Zellkomplex. Zellen mit beiderlei Chromatophoren werden auch weiterhin aufspalten können usw...."
Diese Folgerung führte zur Formulierung der Entmischungshypothese (E. BAUR, 1909, O. RENNER, 1922). Sie geht davon aus, daß in einer Zelle gleichzeitig verschiedene Plastidensorten vorkommen. Daß das tatsächlich zutrifft, konnte aber erst Jahre später durch das Auftreten von "Mischzellen" bewiesen werden.
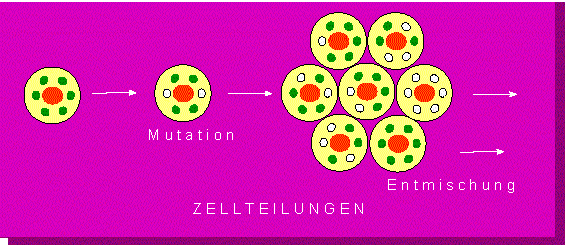
P. MICHAELIS (1955/1956, Max-Planck-Institut für Züchtungsforschung, Köln-Vogelsang) berechnete unter verschiedenen Voraussetzungen die statistische Wahrscheinlichkeit einer Entmischung zweier Plastidensorten. In Zellen höherer Pflanzen kommen größenordnungsmäßig 10-100 Plastiden und etwa 700 Mitochondrien vor. Unter der Annahme, daß zur vollständigen Entwicklung einer einjährigen Pflanze etwa 100 aufeinanderfolgende Zellteilungen erforderlich sind, läßt sich eine praktisch vollständige Entmischung der Plastiden vorhersagen. Für die Mitochondrien trifft diese Aussage nicht mehr zu. Die Entmischung bleibt, bei 700 pro Zelle, über 100 Zellteilungsfolgen unvollständig.
Die Mitochondrien müssen an dieser Stelle ausdrücklich genannt werden, weil auch sie als Erbträger erkannt worden sind. Die Ergebnisse wurden jedoch weniger an Pflanzen als vielmehr an Hefen, anderen Pilzen, Protozoen und tierischen Zellen gewonnen. In den sechziger Jahren wurde schließlich gezeigt, daß sowohl Plastiden als auch Mitochondrien DNS enthalten, welche ihrerseits genetische Information trägt. Vorläufig abschließend kann plasmatische Vererbung nunmehr durch drei Eigenschaften charakterisiert werden
![]() Die Verteilung folgt nicht den MENDELschen Regeln.
Die Verteilung folgt nicht den MENDELschen Regeln.
![]() Reziprok angesetzte Kreuzungen verhalten sich unterschiedlich.
Reziprok angesetzte Kreuzungen verhalten sich unterschiedlich.
![]() Die Erbträger entmischen sich während der Ontogenese
(Embryonalentwicklung).
Die Erbträger entmischen sich während der Ontogenese
(Embryonalentwicklung).
Doch nicht immer beruht eine grün-weiße Scheckung von Blättern oder eine Panaschierung von Blüten auf Entmischung plasmatischer Faktoren, denn:
In vielen Fällen werden grün-weiß-Muster (Chlorosen) durch Viren hervorgerufen; gleiches gilt für "Buntblättrigkeit" mancher Blüten, zum Beispiel der Tulpe. Die Scheckung kann nur durch vegetative Vermehrung oder Pfropfung erhalten werden, weil Viren in fast allen Fällen nicht in die Samen gelangen, die Nachkommenschaft daher gesund (Blätter grün, Blüten einfarbig) ist.
Es gibt eine Vielzahl von regelmäßig strukturierten grün-weiß-Mustern, deren Vererbung nach den MENDELschen Regeln erfolgt, die also durch Gene im Kern determiniert werden.
Im Gegensatz zum eben besprochenen Phänotyp kommen Muster vor, bei denen es so aussieht, als kämen im Verlauf der Ontogenese zahlreiche Umschläge vor. Aus einer weißen Linie entstehen grüne Zellen, die wieder in weiße zurückschlagen können - und umgekehrt. Für diese Erscheinung machte man früher "labile" Gene verantwortlich. Seit einigen Jahren weiß man, daß es tatsächlich "springende" (variable) Gene gibt, die als "Schalter" bestimmte andere Gene ein- und wieder abschalten.
Die Existenz von Genen im Kern, in den Plastiden (und in den Mitochondrien), wirft die Frage nach der Kooperation der einzelnen Genome untereinander auf. Mit anderen Worten: Verträgt sich jeder Kern mit jedem Plasma?
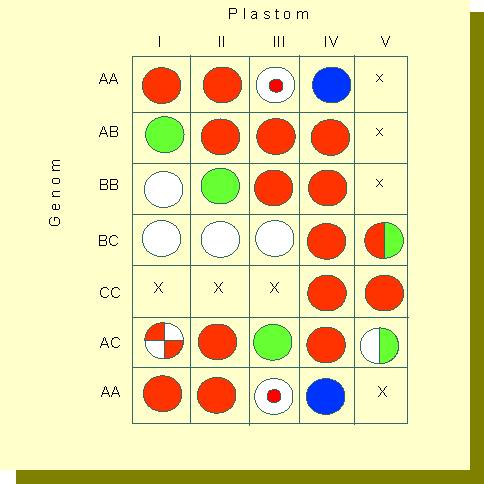
Kombinationsrechteck, das eine Übersicht über die Wirkung bestimmter Genom-Plastom-Kombinationen bei Artkreuzungen mit Arten aus der Untergattung Euoenothera auf der Ergrünungsfähigkeit der Plastiden gibt. Rote Felder: völlige Verträglichkeit (grüne Plastiden), x: letale Kombinationen; weiße Felder: Kombinationen mit nicht-ergrünenden Plastiden; halbiert: Kombinationen mit partiell ergrünenden Plastiden; Felder mit Sektoren oder konzentrischen Kreisen: unterschiedliche Phänotypen in der Nachkommenschaft (Nach W. STUBBE, 1959)
Diese Situation tritt z.B. stets bei Artkreuzungen in Erscheinung, von denen man ja weiß, daß sie gelegentlich zustande kommen, oft aber auch mißlingen. Vielfach kommt es bei Artkreuzungen zu einer "Bastardbleichheit", einer Erscheinung, die O. RENNER in den zwanziger und dreißiger Jahren an Arten der Gattung Oenothera analysierte. In reziproken Kreuzungen der beiden Arten Oenothera hookeri x Oenothera lamarckiana fallen die folgenden Unterschiede ins Auge:
(1) (weiblich) Oe. hookeri x (männlich) Oe. lamarckiana
Fl: Bastarde fast immer grün. Selten traten Pflanzen mit helleren Flecken auf den ersten Blättern auf.
(2) (weiblich) Oe. lamarckiana x (männlich) Oe. hookeri
F1: Bastarde bleich, viele Jungpflanzen gingen zugrunde.
Ein Teil der Bastarde hatte jedoch auf den Keimblättern und den ersten Laubblättern normal grüne Flecken.
RENNER führte die Unterschiede auf Verschiedenheiten im Erbgut der Plastiden zurück, wobei die Plastiden vom Genotyp der Kerne beeinflußt werden. Durch umfangreiche Serien von Kreuzungsexperimenten mit einer Anzahl von Oenothera - Arten wurden in den letzten Jahrzehnten verschiedene Plastidentypen und unterschiedliche Genomkomplexe charakterisiert und die Verträglichkeit der einzelnen Kombinationen untereinander getestet (F. SCHÖTZ, Botanisches Institut der Universität München, W. STUBBE, Botanisches Institut der Universität Düsseldorf). Dabei zeigte sich nach Prüfung von etwa 400 Kombinationen, daß die Plastiden von 14 Wildarten zu 5 Reaktionstypen (I-V) zusammengefaßt werden können. Zu den Unterschieden gehört eine unterschiedliche Vermehrungsgeschwindigkeit der Plastiden. So lassen sich die einzelnen Oenothera-Arten nach abnehmender Vermehrungsrate in folgender Reihe anordnen:
hookeri > bauri > lamarckiana > rubricaulis > suaveolens > biennis > syrticola > parviflora > rubricuspis > ammophila > artrovirens.
Die Vermehrungsgeschwindigkeit wird im wesentlichen von den Plastiden selbst und nicht vom Kern bestimmt. Zum Vergleich können die Eigenschaften der Kerne klassifiziert werden (A-C). Die Ergebnisse der Untersuchungen lassen sich zu einem Schema zusammenstellen, aus dem ersichtlich wird, daß es einen unterschiedlichen Grad an Verträglichkeit zwischen Kern-Plastiden-Kombinationen gibt.
Der Einfluß eines bestimmten Kerngens auf die Ergrünungsfähigkeit der Plastiden des Mais wurde 1946 von M. M. RHOADES (Cold Spring Harbor Laboratory) untersucht. Das Gen Iojap (Ij) verursacht im homozygot rezessiven Zustand eine (regelmäßige) grün-weiß-Streifung der Maisblätter. Zunächst sah es so aus, als würde es sich entsprechend den Mendelschen Regeln verhalten. Detaillierte Kreuzungsexperimente führten jedoch zu folgendem Ergebnis:
(1) (weiblich) grün (IjIj) x (männlich) gestreift (ijij)
F1: einheitlich grün (Ijij)
(2) (weiblich) gestreift (ijij) x (männlich). grün (IjIj)
Fl: vorwiegend grün (Ijij), daneben gestreift (Ijij) und weiß (letal) (Ijij).Trotz unterschiedlicher Phänotypen bleibt der Genotyp stets gleich.
(3) Durch Kreuzung von (weiblich) gestreift (Ijij) x (männlich) grün (Ijij) erhält man folgende Typen (entsprechend einer F2)
grün (IjIj)
gestreift (IjIj)
weiß (letal) (IjIj)
grün (Ijij)
gestreift (Ijij)
weiß (letal) (Ijij).
Die Analyse der Daten läßt den Schluß zu, daß das Gen ij die Bildung abnormaler Chloroplasten induziert, welche anschließend als solche erhalten bleiben und sich unabhängig von der Konstitution des Kerns manifestieren. Deshalb können in der F2 auch in Pflanzen des Genotyps IjIj oder Ijij gestreifte (und weiße) Formen auftreten. Wie entsteht nun aber die Streifung? Warum entstehen durch ijij-Einfluß nicht ausschließlich defekte (= weiß aussehende) Plastiden? Offensichtlich bleibt auch in den ijij-Pflanzen eine Anzahl von Plastiden grün, und es sieht so aus, als könnten gelegentlich auch die farblosen ergrünen. Jedenfalls machen die Ergebnisse deutlich, daß die genetische Information von Kern und Plastiden einander ergänzt und daß die Genome der unterschiedlichen Kompartimente zusammenarbeiten.
Molekulargenetische Untersuchungen der letzten Jahre ergaben, daß es einen perfekt funktionierenden, genau aufeinander eingespielten Informationsfluß zwischen dem Kern und den Plastiden (und den Mitochondrien) gibt und daß die in Chloroplasten und Mitochondrien lokalisierten Gene einen entscheidenden (unersetzlichen) Einfluß auf die Existenz der Zellen haben. In Chloroplasten lokalisierte Gene steuern Reaktionen der Photosynthese, in Mitochondrien lokalisierte den Reaktionsablauf der Atmungskette.
|
|