
Auf J. B. van HELMONT (1577-1644) geht die Beobachtung zurück, daß eine Weide, die fünf Jahre in einem Gefäß gestanden und gehörig gegossen wurde, über einen halben Zentner an Gewicht zunahm, obwohl die Erde im Topf nur zwei Unzen abgenommen hatte. Der britische Naturforscher S. HALES (1677-1761) bemerkte, daß Luft und Licht für die Ernährung der grünen Pflanzen erforderlich seien. Doch erst nachdem man erkannt hatte, daß die Luft aus verschiedenen Gasarten zusammengesetzt ist, begann man, sich mit ihrer Bedeutung für die Pflanzenernährung zu befassen. J. PRIESTLEY (1733-1804), einer der Entdecker des Sauerstoffs, fand 1771, daß grüne Pflanzen Sauerstoff abgeben und damit die Luft verbessern.
Der Genfer Priester J. SENEBIER (1742-1809) bemerkte, daß die Regeneration der Luft auf dem Verbrauch von "fixer Luft" (Kohlensäure) beruht. Bestätigt und erweitert wurden diese Befunde durch Arbeiten (ab 1779) des holländischen Arztes J. INGENHOUSZ (auch INGEN-HOUSZ geschrieben; 1730-1799), der sowohl die Bedeutung des Lichts für die Kohlensäureaufnahme erkannte als auch feststellte, daß der gesamte in Pflanzen enthaltene Kohlenstoff atmosphärischen Ursprungs ist. Neben der lichtabhängigen Kohlensäureaufnahme fand er, daß Pflanzen im Schatten und in der Nacht (in kleinen Mengen) Sauerstoff aufnehmen und dabei Kohlensäure produzieren. Th. des SAUSSURE (1767-1845) aus Genf stellte 1804 fest, daß die Gewichtszunahme von Pflanzen nicht allein auf der Kohlenstoffaufnahme und der Aufnahme von Mineralien beruhen konnte, und folgerte daraus, daß die Zunahme durch Bindung der Bestandteile des Wassers zu erklären sei.
1894 baute T. W. ENGELMANN (1843-1909) aus einem modifizierten Mikroskopkondensor eine Vorrichtung, mit der er Teile photosynthesaktiver Zellen (der Grünalge Spirogyra) durch einen feinen Lichtstrahl beleuchtete, um somit zu klären, welche Komponenten der Zelle als Lichtrezeptoren wirken. Um eine Sauerstoffproduktion zu messen, tauchte er den Spirogyra-Faden in eine bakterienhaltige Suspension. Wurden Teile des Chloroplasten illuminiert, konzentrierten sich die Bakterien in der belichteten (und somit sauerstoffreichen) Zone. Bei Belichtung anderer (farbloser) Bereiche der Zelle blieb die Anlockung der Bakterien aus.
In einer vorangegangenen Arbeit (1882) zerlegte er weißes Licht durch ein Prisma in die Spektralfarben und beleuchtete mit diesem Spektrum einen Faden der Grünalge Cladophora (im Gegensatz zu Spirogyra füllt ihr Chloroplast jede Zelle gleichmäßig aus). Dabei sah er, daß sich die Bakterien vorwiegend an solchen Abschnitten der Zelloberflächen sammelten, die blauem oder rotem Licht ausgesetzt waren. Damit wurde ein erstes Aktionsspektrum (Wirkungsspektrum) der Photosynthese gewonnen. In groben Zügen ähnelt es den Absorptionsspektren von Chlorophyll a und b.
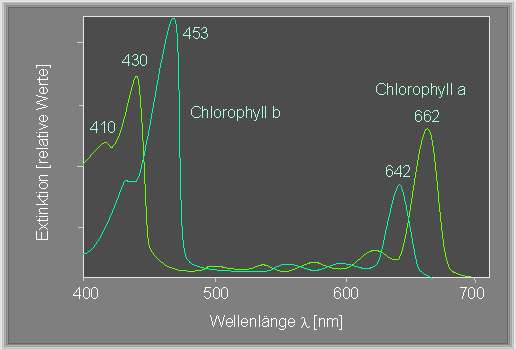
Der Nachweis, daß Chlorophyll an der Photosynthese beteiligt ist, wurde von dem Würzburger Pflanzenphysiologen J. v. SACHS (1832-1897) erbracht. Er - zeigte ferner, daß in Chloroplasten - als Folge photosynthetischer Aktivitäten Stärke gebildet wird.
Diese Befunde standen im Einklang mit dem ersten Hauptsatz der Thermodynamik (Satz von der Erhaltung der Energie), dessen Entdecker J. R. MAYER bereits 1842 postulierte, daß Pflanzen Energie in Form von Licht aufnehmen und sie in eine andere Energieform - in chemische Energie - transformieren. Auf diesen Befunden aufbauend, wurde die Reaktionsgleichung
6 Kohlendioxyd + 6 Wasser > (Chlorophyll) > Glucose + 6 Sauerstoff
J. v. LIEBIG nahm an, der Sauerstoff ginge aus einer Kohlendioxyd-Spaltung hervor. Diese Vorstellung wurde von den bekannten Pflanzenphysiologen des ausgehenden 19. und des beginnenden 20. Jahrhunderts (SACHS, PFEFFER, JOST u.a.) kritiklos übernommen, obwohl M. J. SCHLEIDEN schon 1842 erkannte, daß
- bei der Photosynthese Traubenzucker gebildet wird (damit war er der Realität näher als nach ihm SACHS), und daß
- vermutlich das Wasser gespalten wird. Er schreibt dazu:
"Nun weiß man aber, daß CO2 eine der allerfestesten Verbindungen ist, deren Zersetzung in der Chemie auf keinem Wege gelingt, dagegen ist bekannt, daß H2O eine gar leicht zersetzbare Verbindung ist.... und so erscheint es wahrscheinlich, daß sich mit 12 CO2 die 24 H2 von 24 H2O verbinden."
(aus: Grundzüge der wissenschaftlichen Botanik) (Anmerkung: SCHLEIDENs Reaktionsgleichung enthält alle Reaktionspartner in Zahlen, die doppelt so hoch sind, wie in den heute üblichen Darstellungen. Für Traubenzucker nennt er die Zusammensetzung C12H24O12
Pflanzenphysiologen merkten schon bald, daß die oben dargestellte Reaktionsgleichung eine grobe Vereinfachung ist und daß die Photosynthese aus einer Reihe von Teilabläufen bestehen muß.
F. F. BLACKMAN und G. L. C. MATHGEL (1905, Universität Cambridge/England) gehörten zu den ersten, die diesen Fragenkreis systematisch angingen. Sie kultivierten Pflanzen unter unterschiedlichen, aber kontrollierten Kohlendioxyd-Konzentrationen, unterschiedlichen Lichtintensitäten und unterschiedlichen Temperaturen und registrierten die Auswirkungen dieser externen Parameter auf die Photosyntheserate. Dabei ergaben sich zwei entscheidende Aspekte. Unter Starklichtbedingungen (bei ausreichender Lichtmenge) und limitierenden Kohlendioxyd-Konzentrationen war die Photosyntheserate temperaturabhängig. Das verdeutlichte, daß die Kohlendioxyd-Verwertung (Fixierung) auf normalen temperaturabhängigen biochemischen Reaktionen beruht. Bei Kohlendioxyd-Überschuß und nicht ausreichender Lichtmenge wurde keine Temperaturabhängigkeit gefunden. Das wiederum weist darauf hin, daß die lichtinduzierten Reaktionen temperaturunabhängig sind. Diese Aussage gilt ganz allgemein für photochemische Reaktionen.
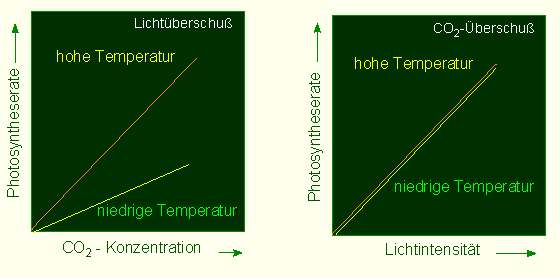
O. WARBURG (Kaiser-Wilhelm-Institut [später Max-Planck-Institut] für Zellphysiologie in Berlin-Dahlem) führte 1925 BLACKMANs Ergebnisse auf die Existenz von zwei Reaktionsklassen, die Lichtreaktionen und die Dunkelreaktionen, zurück.
In den dreißiger Jahren analysierte C. B. van NIEL (Stanford University) die Photosynthese einer Reihe von Purpurbakterien, die dafür außer dem Kohlendioxyd Schwefelwasserstoff benötigen. Als Reaktionsgleichung konnte van NIEL
6 CO2 + 12 H2S > (Licht) > C6Hl2O6 + 12 S + 6 H2O
ermitteln. Hiervon extrapolierend, stellte er eine generelle Gleichung für alle Photosyntheseprozesse auf:
CO2 + 2 H2X > (Licht) > (CH2O) + H2O + 2 X
Demnach wäre eine Photosynthese eine Redoxreaktion mit H2X als Elektronendonator (der oxydierbaren Substanz). Im Falle grüner Pflanzen wäre es das H2O, und das wiederum heißt, daß nicht das Kohlendioxyd, sondern das Wasser gespalten wird.
Ein erster experimenteller Beweis dafür, daß der Sauerstoff bei der Photosynthese grüner Pflanzen tatsächlich aus der Spaltung des Wassers stammt, erfolgte 1937 durch den britischen Physiologen R. HILL. Er stellte fest, daß isolierte Chloroplasten in Anwesenheit zugesetzter (unnatürlicher) reduzierender Verbindungen (z.B. Eisenoxalat, Ferricyanid, Benzochinon) nach Belichtung Sauerstoff freisetzen. Die Reaktion ging als HILL-Reaktion in die Literatur ein:
2 H2O + 2 A > (Licht, Chloroplasten) > 2 AH2 + O2
(A = Elektronenakzeptor) Wenn A = FeIII , erhält man
2 H2O + 4 FeIII > (Licht, Chloroplasten) > 4 FeII + O2 + 4 H+
Der Ablauf ist mit einer photolytischen Wasserspaltung verbunden, die der Reduktion des FeIII vorangeht.
4 H2O > (Licht, Chloroplasen) > 4 H+ + 4 OH-
| Sauerstoff auch in Kohlendioxyd-Abwesenheit freigesetzt werden kann, | |
| der gebildete Sauerstoff aus der Wasserspaltung stammt | |
| und daß schließlich in isolierten Chloroplasten - zumindest Teilschritte - der Photosynthese ablaufen können. |
Die Aussage, daß der bei der Photosynthese produzierte Sauerstoff ausschließlich aus der Spaltung des Wassers stamme, wurde - nach Einführung der Isotopentechniken in die Biochemie - 1941 von S. M. R RUBEN, M. RANDALL, M. KAMEN und J. L. HYDE bekräftigt, als sie zeigen konnten, daß eine Suspension von Chlorella-Zellen nach Belichtung aus H218O 18O2 freisetzt. Unmittelbar danach bestätigten S. M. RUBEN und Mitarbeiter O. WARBURGs Postulat, daß die Fixierung des Kohlendioxyds zwar energieverbrauchend, aber lichtunabhängig ist. E. RACKER (Cornell University, Ithaca, N.Y.) ergänzte die Aussage und fand, daß die Lichtenergie durch Zugabe energiereicher Verbindungen ersetzt werden kann.
|
|