
Wie im historischen Überblick dargelegt, wird für die Photosynthese Licht benötigt, und wie schon die Ergebnisse von ENGELMANN und SACHS zeigten, wird es primär von Chlorophyll absorbiert. An anderer Stelle haben wir gesehen, daß es bei Pflanzen zwei Typen von Chlorophyll (a und b) gibt und daß sich jedes durch ein charakteristisches Absorptionsspektrum auszeichnet.
Das Aktionsspektrum der Photosynthese ähnelt den Absorptionsspektren, ist mit diesen jedoch nicht identisch. Das bedeutet, daß es neben den Chlorophyllen weitere Photorezeptoren (akzessorische Pigmente) geben muß.
Nachdem wir im vorangegangenen Abschnitt den Verbleib des Kohlendioxyds verfolgt haben, wäre jetzt die Frage zu klären: Welche Reaktionen induziert das Licht und wie wird Lichtenergie in chemische Energie umgesetzt? Mit anderen Worten: Wie werden ATP und NADPH2 erzeugt?
Die Dunkelreaktionen hatten CALVIN und Mitarbeiter an intakten, aktiven Zellen studiert. Zur Klärung der Lichtreaktionen genügte dieser Ansatz nicht. Die Ergebnisse führten zu kontroversen Aussagen, und es mußten daher zunächst Verfahren entwickelt und verfeinert werden, um Zellfraktionen (isolierte Chloroplasten) in aktivem Zustand zu gewinnen.
Nachdem der Gebrauch solcher Fraktionen zur Routine wurde, konnten 1951 drei Arbeitsgruppen gleichzeitig und unabhängig voneinander zeigen, daß isolierte Chloroplasten bei Belichtung NADP zu NADPH2 reduzieren können [W. VISHNIAC und S. OCHOA (Rockefeller Institute, New York), L. J. TOLMACH (University of Chicago) und D. I. ARNON (University of California, Berkeley)]. Kurz danach (1954) fanden ARNON und Mitarbeiter, daß auch die Produktion von ATP lichtabhängig ist und daß sowohl ATP als auch NADPH2 gleichzeitig gebildet werden können. Beide Verbindungen entstehen aus Vorstufen, die schon vor Beginn der Photosynthese in den Chloroplasten enthalten waren, da jene während des Experiments von jeglicher Zufuhr externer Metaboliten abgeschnitten wurden. Licht (Photonen) war demnach die einzige gebotene Energiequelle. Es stellte sich heraus, daß für die ATP-Bildung Sauerstoff weder benötigt noch dabei freigesetzt wurde, so daß die Gleichung
n ADP + n Pi > (Photonen) > n ATP
formuliert werden konnte. Man bezeichnet diesen Vorgang als Photophosphorylierung. Die gleiche Erscheinung wurde auch bei photosynthetisierenden Bakterien und bei Blaualgen nachgewiesen und konnte daher als ein allgemeines Phänomen von Photosyntheseprozessen angesprochen werden.
Nachdem sichergestellt war, daß ATP gebildet wird, mußte man sich fragen, wie es geschieht. Es schien unwahrscheinlich, daß das Licht die ATP-Bildung direkt bewirkt, doch mußte seine Wirkung der ATP-Bildung vorangehen, so daß man das Konzept eines lichtinduzierten Elektronenflusses entwickelte. Die Hypothese eines Elektronenflusses sieht vor, daß ein Chlorophyllmolekül ein Lichtquant (Photon) absorbiert, dadurch in einen angeregten Zustand übergeht, d.h. daß ein Elektron auf ein höheres Energieniveau gebracht wird.
Dieses energiereiche Elektron wird dann auf ein benachbartes Elektronenakzeptormolekül übertragen, das seinerseits über ein starkes elektronegatives Redoxpotential verfügt. Die Überführung des Elektrons vom angeregten Chlorophyllmolekül auf den (ersten) Akzeptor beendet bereits die photochemische Phase der Photosynthese. Ihr entscheidender Aspekt liegt in der Tatsache begründet, daß ein Photonenfluß in einen Elektronenfluß transformiert wird.
Sobald eine stark elektronennegative (reduzierende) Substanz gebildet worden ist, kann ein Elektronenfluß auf weitere Elektronenakzeptoren (mit weniger negativen Redoxpotentialen) erfolgen, wobei chemische Energie frei wird, die nunmehr zur Photophosphorylierung genutzt werden kann. Es gab bereits in den fünfziger Jahren sichere Hinweise darauf, daß die Chloroplasten-Cytochrome an dieser Elektronentransportkette beteiligt sind. Man konnte auch zeigen, daß das Elektron am Ende des Prozesses wieder zum Chlorophyll zurückkehrt, so daß dessen ursprünglicher Zustand wiederhergestellt wurde und somit die Bedingung einer Katalyse erfüllt war. Dieser Vorgang ist als zyklische Phosphorylierung bekannt geworden (D. I, ARNON, 1959).
Ein solcher zyklischer Elektronenfluß, der durch Licht angetrieben wird und chemische Energie freisetzt, die zur Bildung einer energiereichen Bindung in ATP genutzt wird, ist einzigartig und zeichnet photosynthetisierende Zellen aus.
Konzept der zyklischen Photophosphorylierung (links); rechts: das ursprüngliche Konzept der nichtzyklischen Photphosphorylierung (Nach D. I. ARNON, 1971)
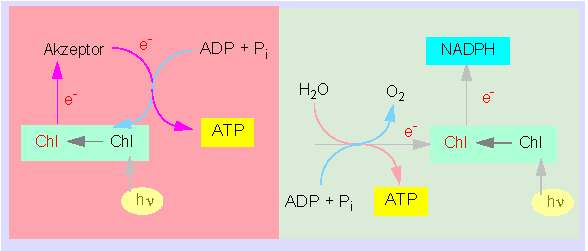
Jetzt fehlt noch eine Erklärung zur Photoreduktion von NADP in Chloroplasten. Wiederum waren es ARNON und Mitarbeiter, die 1957 einen zweiten Typ einer Photophosphorylierung entdeckten und damit einen direkten experimentellen Beweis für die Kopplung zwischen der Photoreduktion des NADP und der Synthese von ATP erbrachten. Im Gegensatz zur zyklischen Photophosphorylierung ist die ATP-Bildung hier stöchiometrisch mit einem lichtbetriebenen Transfer von Elektronen aus dem Wasser auf NADP und einer Sauerstoffproduktion gekoppelt. Die ATP-Bildung im Gesamtsystem steigerte die Reduktionsrate von NADP, dies wiederum wies darauf hin, daß dieser Prozeß und die zyklische Photophosphorylierung eng miteinander verbunden sind. Da Elektronen vom Chlorophyll irreversibel auf NADP übertragen wurden, benötigte man Ersatz, und diese Elektronen entstammen der Spaltung des Wassers. Man spricht daher hier, da gleichzeitig ATP gebildet wird, von nichtzyklischer Photophosphorylierung. Eine Schlüsselrolle spielt dabei das Ferredoxin (ein hämfreies Eisen-Schwefel-Protein). Sein Reduktionspotential ist weit negativer als das von NADP, so daß ein Elektronenfluß von ihm auf NADP nahelag. Die Reduktion des NADP erwies sich als eine Dreischrittreaktion
- eine photochemische Reduktion des Ferredoxins, der zwei "Dunkel"-Schritte folgten.
- Reoxydierung von Ferredoxin durch eine Ferredoxin-NADP-Reduktase (ein Flavoprotein).
- eine Reoxydierung der Ferredoxin-NADP-Reduktase durch NADP.
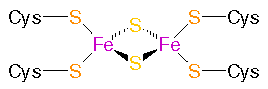
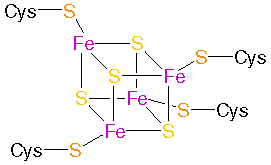
Ferredoxin: Eisen-Schwefel-Komplex: links: Ferredoxin aus Pflanzen (Fe2S2 Proteine - Plant�type ferredoxins) , rechts: Ferredoxin aus Bakterien
(Fe4S4 proteins - Bacterial�type ferredoxins)
aus: PROMISE - The Prosthetic groups and Metal Ions in Protein Active Sites Database
Das, was also ursprünglich als eine Photoreduktion des NADP angesehen wurde, ist demnach eine Elektronentransportkette, die über Ferredoxin und eine Flavinkomponente zu NADP läuft.
Die herausragende Stellung des Ferredoxins wurde noch weiter gefestigt, als man feststellte, daß bei der Reaktion in stöchiometrischen Mengen O2 und ATP entstanden, so daß man die nichtzyklische Photophosphorylierung wie folgt darstellen konnte:
4 Ferredoxin (oxyd.) + 2 ADP + 2 Pi + 2 H2O > (Photonen) > 4 Ferredoxin (red.) + 2 ATP + O2 + 4 H+ .
Nach diesen Befunden stellte sich die Frage, wie diese Reaktion mit der eingangs besprochenen zyklischen Photophosphorylierung zusammenhängt. Durch eine Serie von Experimenten, bei denen spezifische Inhibitoren eingesetzt wurden, wurde sichergestellt, daß das Ferredoxin auch dort ein Glied in der Reaktionskette ist.
Soweit die chemischen Daten. Um sie zu deuten, mußte man Genaueres über die primäre Wirkung des eingestrahlten Lichts und über die Bedeutung des Chlorophylls wissen. Auch diese Probleme haben eine lange Vorgeschichte.
1932 haben R. EMERSON und U. ARNOLD von der University of Illinois in Urbana Chlorella-Zellen einer Abfolge von extrem kurzen Lichtblitzen ausgesetzt. Das Experiment sollte klären, wie viele Chlorophyllmoleküle benötigt werden, um ein Lichtquant (Photon) zur Produktion eines Sauersoff-Moleküls zu nutzen. Das Ergebnis lautete: Es werden mehrere hundert Chlorophyllmoleküle gebraucht, und das wiederum besagt, daß sie nicht alle gleichwertig sind. Der überwiegende Teil wirkt als Lichtfalle (oder Antenne), die dafür sorgt, daß ein Photon zu einem Reaktionszentrum kanalisiert (transferiert) wird, in dem schließlich an einem besonders exponierten Chlorophyllmolekül Lichtenergie in chemische Energie umgewandelt wird. H. GAFFRON nannte diesen Komplex aus hunderten von Chlorophyllmolekülen, an dem auch andere Pigmente (Carotine, Carotinoide, Xanthophylle u.a.) und Proteine beteiligt sind, eine photosynthetische Einheit.
Ganz offensichtlich führt diese Aggregation von Pigmenten zu einer besonders effizienten Ausnutzung des eingestrahlten Lichts. Dennoch geht ein nicht unbeträchtlicher Teil der eingestrahlten Energie verloren. Er erreicht nicht das Reaktionszentrum und wird in Form von Wärme oder Licht (rote Eigenfluoreszenz des Chlorophylls) wieder abgestrahlt.
Betrachtet man das Aktionsspektrum der Photosynthese, fällt auf, daß die Effizienz von Licht der Wellenlängen lambda > 680 nm stark abfällt, obwohl Chlorophyll a in dem Bereich noch eine merkliche Absorption aufweist. R. EMERSON (1957) fand, daß Licht der Wellenlänge von lambda = 700 (710) nm bei gleichzeitiger Einwirkung von Licht der Wellenlänge lambda = 680 nm (oder darunter) die Photosyntheserate drastisch steigert.
Bei unabhängiger oder aufeinanderfolgender Applikation der beiden Lichtqualitäten unterbleibt der Steigerungseffekt (EMERSON-Effekt). Hieraus schloß er, daß es zwei photochemische Ereignisse geben müsse, die zwar mit unterschiedlichen Pigmentsystemen (Lichtrezeptoren) arbeiten, aber miteinander kooperieren. Wir nennen sie heute (einem Vorschlag von L. N. M. DUYSENS folgend):
| Photosystem I (PS I). Es benötigt Licht längerer Wellenlängen (lambda > 700 nm). | |
| Photosystem II (PS II). Es arbeitet mit Licht kürzerer Wellenlängen (lambda < 680 nm und darunter). |
Das Verhältnis Chlorophyll a zu b im PS I ist höher als im PS II. Zu klären bleibt, wie die beiden Systeme miteinander kooperieren und wie sie mit der besprochenen ATP- und NADPH2-Bildung zusammenhängen.
ARNON und Mitarbeiter wiesen nach, daß sie in Serie geschaltet sind und daß beide benötigt werden, um alle Effekte, die man als photosynthesespezifisch erkannt hatte, zu erklären. Nur bei einigen photosynthetisierenden Bakterien, bei deren Photosynthese kein Sauerstoff produziert wird, fehlt das Photosystem II. Schon dieser Befund legt die Annahme nahe. daß die Wasserspaltung an das Photosystem II gekoppelt ist und daß das Photosystem I in der Evolution vor dem Photosystem II entstanden ist.
Das Reaktionszentrum eines jeden Photosystems wird durch je ein Molekül Chlorophyll a repräsentiert (P700 für PS I und P680 für PS II; die Abkürzung P bedeutet hier Pigment).
Die Absorption eines Photons durch P680 (im Grundzustand mit positivem Redoxpotential: +0,8 V) überführt es in seinen angeregten Zustand (Redoxpotential: 0,0 V) und bewirkt die Bildung einer stark oxydierenden Komponente (Z+) und einer schwach reduzierenden (Q-). Z+ entzieht dem Wasser Elektronen, wobei O2 und Protonen frei werden:
4 Z+ + 2 H2O > 4 Z + 4 H+ + O2
Die reduzierende Komponente (ein membrangebundenes Plastochinon) schleust das Elektron in eine Elektronentransportkette ein, in deren Verlauf es an Energie verliert, von der ein Teil zur ATP-Bildung genutzt wird. Das Elektron kehrt nicht zu seinem Ausgangspunkt (dem Chlorophyllmolekül P680) zurück, sondern wird auf ein Chlorophyllmolekül des Photosystems I (P700) übertragen, wodurch beide Systeme miteinander gekoppelt werden.
Unter Absorption eines weiteren Photons wird jenes (P700, Redoxpotential im Grundzustand: + 0,4 - + 0,5 V) angeregt. Es überträgt ein Elektron auf membrangebundenes Ferredoxin (P430), von dem aus es auf gelöstes Ferredoxin weitergeleitet wird. Die folgenden Schritte sind uns bekannt.
Stöchiometrisch betrachtet ist die obige Darstellung noch nicht vollständig, denn Ferredoxin überträgt jeweils nur ein Elektron, während zur NADPH2-Bildung zwei Elektronen benötigt werden. Richtig muß es demnach heißen:
2 Ferredoxin (red.) + 2 H+ + NADP+ > 2 Ferredoxin (oxyd.) + NADPH2
Wir könnten nunmehr zusammenfassend sagen, daß die Lichtenergie für einen Elektronenfluß von Wasser auf NADPH2 bei gleichzeitiger ATP-Bildung eingesetzt wird (Z-Schema).
Bei der Behandlung dieser Elektronentransportketten wurde die eingangs besprochene zyklische Photophosphorylierung ausgeklammert. In der Tat erwies sie sich als eine Art Parallelschaltung, die immer dann zum Tragen kommt, wenn ausreichend NADPH2 vorhanden ist, zusätzlich jedoch ATP benötigt wird. An der zyklischen Photophosphorylierung ist nur das Photosystem I beteiligt.
|
|