Koleoptilen von Gräsern (z.B. die des Hafers, die Avena-Koleoptile) sind beliebte Versuchsobjekte der Pflanzenphysiologie. Bei Belichtung einer wachsenden Koleoptile von der Seite, wächst diese in Richtung des einfallenden Lichts weiter. Dieser Vorgang ist bei Pflanzen verbreitet und als Phototropismus bekannt. In seinem Werk über "Das Bewegungsvermögen der Pflanzen" sprach C. DARWIN 1880 der Koleoptilspitze eine entscheidende Rolle bei der Erkennung des Lichtreizes zu und beobachtete, daß die eigentliche Krümmung in einer Zone unterhalb der Spitze erfolgte. Folglich schloß er, daß es eine "Reizleitung" im Gewebe geben müsse. Pflanzenanatomische Untersuchungen ergaben, daß das Wachstum zum Licht auf einem Streckungswachstum von Zellen auf der dem Licht abgewandten Seite beruht. Wird die Koleoptilspitze entfernt, unterbleibt die phototrope Reaktion. Durch Wiederaufsetzen der abgeschnittenen Spitze kann sie jedoch erneut induziert werden. Der Versuch weist auf die Existenz einer Substanz hin, die polar (von oben nach unten: basalwärts, basipetal) wandert und das Streckungswachstum verursacht.
Der dänische Botaniker P. BOYSEN-JENSEN unterbrach den vermeintlichen Stofftransport durch Einsetzen eines Glimmerplättchens an der lichtabgewandten Seite und trennte damit die Koleoptilspitze und das darunterliegende Gewebe (1913). Das Ergebnis: Die phototrope Reaktion unterblieb. Folglich gibt es keinen Transport eines Effektors "um die Ecke herum". In Kontrollversuchen, in denen das Glimmerplättchen an der belichteten Stelle oder senkrecht zur Achse eingesetzt wurde, blieb die Reaktion erhalten.
Ende der zwanziger Jahre wurde die stoffliche Natur des Effektors durch den holländischen Pflanzenphysiologen F. WENT endgültig sichergestellt. Er ging davon aus, daß eine wandernde Substanz auch in einen Gelatineblock einwandern müsse, und setzte daher abgeschnittene Koleoptilspitzen mit der Schnittstelle auf kleine Gelatineblöcke. Einige Zeit danach entfernte er die Spitzen und setzte die Blöcke, von denen er annahm, daß sie den Effektor aufgenommen hatten, auf dekapitierte Koleoptilen.
Über die Durchführung des entscheidenden Experiments schreibt er:
"When I removed the tip after an hour and placed the gelatine cube on one side of the seedling, nothing happened at first. But in the course of the night, the stump started to curve away from the gelatine block. It had acquired the capacity of the stem tip to grow! At 3:00 A.M. on April 17, 1926"
(zitiert aus F. B. SALISBURY, C. W. ROSS: Plant Physiology. Belmont/Cal: Wadsworth Publ. Comp. 1978, 2. Aufl.)
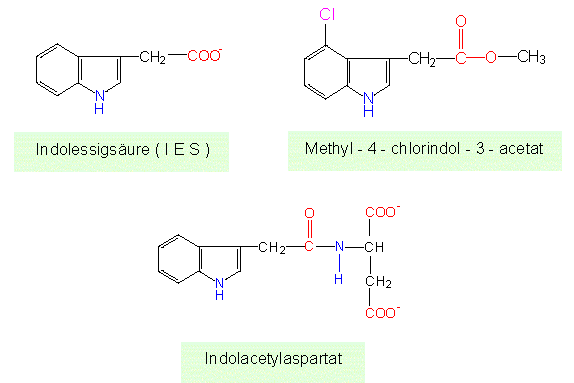
Went nannte den Effektor Auxin (oder Wuchsstoff). Chemisch erwies er sich als Indol-3-Essigsäure (IES), und wie die Formel vermuten läßt, ist er ein Tryptophanderivat.
In Zellen liegt er in Konzentrationen von 10-8 -10-6 Mol/l vor. IES und ähnliche Verbindungen (Auxin ist ein Sammelname) sind, wie wir heute wissen, in grünen Pflanzen (und in Pilzen) weitverbreitet. So wurden in unreifen Erbsensamen (und Samen anderer Pflanzen) z.B. Methyl-4-Chloroindol-3-Acetat, 4-Chloroindol-3-Acetat und Indolacetylaspartat gefunden. Oft sind die Auxine glykosyliert oder an Proteine gebunden.
Die Raten von Auf- und Abbau sowie die Modifizierbarkeit von IES determinieren die Konzentration der physiologisch aktiven Form in der Zelle. In den letzten Jahren sind eine Anzahl von IES-Analogen synthetisiert und auf ihre Hormonwirksamkeit hin überprüft worden, wobei sich herausstellte, daß eine hormonell wirksame Substanz drei Strukturmerkmale aufweisen muß:
- Im Molekül muß ein Ringsystem mit mindestens einer Doppelbindung vorliegen.
- Der Doppelbindung benachbart muß eine Seitenkette vorhanden sein.
- Es wird eine Karboxylgruppe benötigt, die durch ein oder zwei C-Atome vom Ringsystem getrennt ist.
Diese Bedingungen führten zu einigen Anhaltspunkten über die Struktur der Bindungsstelle(n). Demnach muß es am Rezeptormolekül zwei voneinander getrennte Kontaktstellen geben.
Die Ausbreitung von Auxin in pflanzlichen Geweben erfolgt nach einigen klar erkennbaren Regeln, die darauf schließen lassen, daß ein aktiver und zugleich polarer Transport vorliegt. Das heißt, daß es, außer dem oder den Rezeptoren, IES-spezifische Carrier geben muß. Mindestens sechs Gründe sprechen dafür:
- Der Transport erfolgt stets gerichtet, er ist polarisiert.
- Die Transportgeschwindigkeit ist höher, als man aufgrund einfacher Diffusion erwarten würde.
- Der Transport kann gegen ein Konzentrationsgefälle erfolgen.
- Der Transport ist energieabhängig, Abwesenheit von Sauerstoff reduziert ihn drastisch.
- Das Transportsystem ist substratspezifisch. Es transportiert bestimmte Auxinmoleküle, wie z.B. IES und die Naphthylessigsäure schneller als z.B. 2,4 Dichlorphenoxyessigsäure.
- Das Transportsystem kann durch spezifische Inhibitoren blockiert werden.
Welche biologische Bedeutung kommt den Auxinen (IES und vergleichbaren Substanzen) zu? In niederer Konzentration fördern sie das Streckungswachstum von Koleoptilen, der Sproßachsen und der Wurzel. In hohen Konzentrationen werden Wurzel- und Sproßwachstum gehemmt. Der Grund hierfür liegt in einer Produktionsförderung von Äthylen, einem gasförmigen Kohlenwasserstoff, der ebenfalls als Phytohormon erkannt worden ist; eine seiner Eigenschaften ist die Hemmung des Streckungswachstums. Auxine wirken an der Differenzierung von Leitbündeln mit, sie kontrollieren den Blattfall (Abscission), induzieren beta-1,4-Gluconasen in Erbsenwurzeln und fördern das Aufbrechen von Baumknospen sowie das rasche Wachstum junger Triebe. Sie steigern die Zellteilungsrate im Kambium und stimulieren damit das sekundäre Dickenwachstum; weiter bewirken sie das Auswachsen eines Ovars zu einer Frucht und sind für die Ausprägung und Aufrechterhaltung einer Apikaldominanz verantwortlich.
Sie fördern die Plasmaströmung, steigern die Plastizität der Zellwand und verursachen einen Protonenefflux aus der Zelle. Diese Liste an Aktivitäten ist bei weitem nicht vollständig, soll aber andeuten, wie vielseitig Auxinwirkungen sein können.
Zum Schluß noch die Frage nach einem möglichen Wirkungsmechanismus: Wie eingangs angedeutet, lassen die aufgeführten Beispiele physiologischer Effekte darauf schließen, daß die Auxine auf mehrere verschiedene Primärprozesse in der Zelle einwirken. Es gibt experimentelle Belege dafür, daß sie
- die Transkriptionsrate steigern
- die Aktivität bestimmter Enzyme steuern, und
- Einfluß auf Ionenpumpen in der Membran haben.
Zur Klärung des Einflusses auf Membranen sind Modellstudien mit isolierten Membranvesikeln (z.B. mit isolierten Vakuolen) durchgeführt worden. An Plasmamembranvesikeln wurde nachgewiesen, daß es eine pH- und elektropotentialabhängige Akkumulation von Auxin gibt. Der Auxintransport durch die Membran ist gerichtet. Auxin bindet spezifisch an Tonoplasten, und es beeinflußt (in vitro) die Freisetzung von Calciumionen aus Vakuolen. Für den Import und für den Export sind unterschiedliche Carrier nachgewiesen worden, ein Protonencarrier (S), der einen Symport bewirkt und ein Auxin-Anionen-Carrier (AC), ein aktiver Antiport-Carrier. Darüberhinaus ließ sich zeigen, daß diese Auxincarrier in den Membranen asymmetrisch verteilt sind.
|
|