
Der Phototropismus ist eine durch einen Lichtreiz hervorgerufene Wachstumsbewegung. Wachstum auf eine Lichtquelle zu wird als positiver, von der Lichtquelle weg als negativer Phototropismus bezeichnet. Sproßspitzen sind meistens positiv, Wurzelspitzen in der Regel negativ phototrop.
Ursprünglich, auch noch bei J. v. SACHS, wurde der Phototropismus Heliotropismus genannt, weil die Pflanze der Sonne zustrebt, doch erschien ein Namenswechsel sinnvoll, als klar wurde, daß Pflanzen auch auf künstliche Lichtquellen reagieren (W. PFEFFER). Vorweg sei noch ein anderes Problem gestreift: Reagiert die Pflanze auf Licht, oder wächst sie der Luft zu? Dazu ein Zitat von A. de CANDOLLE (1834/38):
"Gewöhnlich sagen die Gärtner und Landwirte, daß die Pflanzen sich nach der freien Luft hinziehen, allein TESSIER hat die Unrichtigkeit dieser Erklärung durch einen einfachen Versuch nachgewiesen: Er brachte lebende Pflanzen in einen Keller mit zwei Öffnungen, von der einen Seite gab ein Glasfenster Licht und keine Luft, von der anderen führte ein Luftloch, das in einen geräumigen dunklen Wagenschauer mündete, Luft, aber kein Licht zu. Alle Pflanzen richteten sich zum Glasfenster."
A. de CANDOLLE bemerkte bereits 1809, daß das Streben zum Licht auf ungleichem Wachstum gegenüberliegender Seiten eines Organs beruht. Die belichtete Seite wächst langsamer als die unbelichtete. J. v. SACHS entdeckte die Bedeutung der Lichtqualität (Wellenlängenabhängigkeit) für die Auslösung der phototropen Reaktion. Blaues, violettes und ultraviolettes Licht zusammen wirken ebenso stark wie Weißlicht. Auch nach Wegfall des UV-Anteils bleibt die Wirkung erhalten. Rotes, gelbes oder grünes Licht ist bei den meisten Pflanzen wirkungslos. Bei einigen Farnprothallien hingegen verursacht gerade rotes Licht eine phototrope Reaktion.
Zwischen der Lichtmenge und der phototropen Reaktion besteht ein meßbarer Zusammenhang, der unter den Stichworten Reizmengengesetz, Produktenregel oder Reziprozitätsgesetz in die Literatur eingegangen ist (FRÖSCHEL und BLAAUW 1908, BLAAUW 1909). Das Reizmengengesetz besagt, daß das Produkt aus Zeit und Intensität, und damit die Energiemenge des eingestrahlten Lichts, das Maß für die Reizstärke sei. Demnach ist es belanglos, ob ein Lichtreiz niederer Intensität über einen längeren Zeitraum hinweg oder ein Reiz hoher Intensität kurzzeitig geboten wird. Wie wir heute aber auch wissen, gilt das Reizmengengesetz nur in sehr engen Grenzen, denn einmal muß eine Mindestlichtmenge vorhanden sein, um die Reaktion überhaupt auszulösen (Schwellenwert), zum anderen hat es sich gezeigt, daß Steigerung der Lichtenergie keineswegs nur zu einer Erhöhung der phototropen Reaktion führt, sondern im Gegenteil, die positive Reaktion sogar unterdrücken kann. Bei fortgesetzter Steigerung hingegen erfolgt erneut eine positive Reaktion, die ein neues Maximum durchläuft, um wieder abzufallen und dann wieder zu steigen (= 1., 2. und 3. positive phototrope Krümmung.
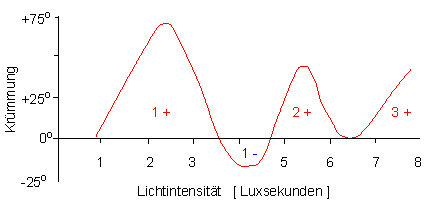
Dosis-Effekt-Kurve des Phototropismus etiolierter Avena-Koleoptilen (schematisierte Darstellung). Die Lichtdosis (Weißlicht) wird in Luxsekunden angegeben; auf der Ordinate ist die positive, bzw. negative Krümmung aufgetragen. Die Reaktionsbereiche der 1., 2. und 3. positiven, sowie der negativen Krümmung sind in der Kurve vermerkt (Nach H. MOHR und P. SCHOPFER, 1978 und H. G. duBUY und E. NUERNBERGK, 1934)
Positiver und negativer Phototropismus im gleichen Gewebe sind nicht auf die Avena-Koleoptilen beschränkt. So wird er z.B. auch unter natürlichen Lichtbedingungen bei Keimlingen der tropischen Araceenart Monstera gigantea beobachtet. Bei geringen Lichtmengen (geringem Photonenfluß) reagieren sie positiv, bei zu hohen negativ phototrop. Unterschiede zu Avena liegen primär in den Reizschwellen, bei denen eine Reaktion in die entgegengesetzte umschlägt.
Phototrope Reaktionen sind für wachsende Gewebe charakteristisch. In ausgewachsenen, ausdifferenzierten Pflanzenteilen sind sie weit schwerer nachweisbar. Das liegt einmal am Verlust der Plastizität der meisten Zellen, zum anderen an der Ausbildung weitgehend starrer Festigungselemente, die jeder Deformation eines Gewebes einen mechanischen Widerstand entgegensetzen. Gerade in der Sproßachse liegen sie meist peripher und verleihen ihr eine besonders hohe Stabilität.
Wie aus dem Dargelegten ersichtlich, haben wir für jeden der drei Reaktionsabschnitte Reizperzeption, Reizweiterleitung, Mechanismus der Krümmungsreaktion, experimentelle Daten, doch bleibt nach wie vor unverstanden, wie diese Glieder miteinander verknüpft sind. Wir wissen nicht, wie der Lichtrezeptor (nach Stimulation durch Licht) die Synthese eines informationstragenden Moleküls (des Phytohormons Auxin) beeinflußt; wir wissen nicht, wodurch die Entscheidung positive oder negative phototrope Reaktion fällt, und wir wissen nur in Ansätzen, wie das Auxin im Bereich der Wachstumszone die Streckung der Zellwand fördern könnte.
In der Lehrbuchliteratur wird als weiteres Beispiel die phototrope Krümmung des Sporangienträgers von Phycomyces genannt. In der Tat kennt man den Mechanismus (vor allem der Lichtperzeption) besser als z.B. bei der Avena-Koleoptile. Es entfällt das Problem der intrazellulären Informationsübertragung. Es stellt sich aber die Frage, wie innerhalb einer Zelle gegenüberliegende Zellwände ungleich schnell wachsen können. Mit Turgoränderungen ist uns hier nicht gedient.
|
|