

The spatial arrangements of vascular bundles is explained by the theory of steles. The term stele sums up the vascular system, associated tissues and the enclosed pith. Different types of steles, like the protostele, the plectostele or the siphonostele exist. Monocot steles are a highly specialized exception.
The hitherto expounded explanations about the structure of vascular bundles are based upon the evaluations of cross-sections of roots and shoots. As we have seen, these preparations are suitable for the depiction of the position and the organization of the vascular bundles (location of the xylem, phloem, cambium, etc.), but they give no hints as to the spatial arrangement and the development of the vascular bundle system. It is rather difficult to get a clear picture here. Serial sections and the derived reconstructions of the vascular bundles' shapes provide a necessary, but very time-consuming method. Another possibility is given by the maceration of the stem, a controlled rotting leading to the assumption that the vascular bundles are the most robust tissues and will therefore survive the maceration procedure rather unimpeded. Despite methodological adversities, the mentioned methods have been used successfully. Today, we know the vascular bundles' spatial arrangements of manifold plants from the most diverse classes; additional information has been gained by the evaluation of fossils.
A first comprehensive, richly depicted survey of the spatial arrangement of vascular bundles can be found in the"Vergleichende Anatomie der Vegetationsorgane der Phanerogamen und der Farne" (Comparative Anatomy of the Organs of Vegetation of Phanerogames And Ferns), published by the plant anatomist A. de BARY from Straßburg in 1877. He recognized the basic patterns, upon which the organization of the vascular bundles of the individual plant groups are founded. Today, it is common to speak of the theory of steles when dealing with the spatial arrangement and development of vascular bundles.
The stele concept goes back to P. van TIEGHEM, professor at the Musée d'Histoire Naturelle de Paris and his pupil H. DOULIOT (1886). Their proposals were went over and modified many times. The terminology suggested by van TIEGHEM is nowadays only of historical interest.
There was, especially at the turn of the 20th century, no well-known morphologist, who was not concerned with this matter. Considerable are particularly E. STRASBURGER (professor of botany at the Universität Bonn) with his work "Über den Bau und die Vorrichtungen der Leitbahnen in der Pflanze" (About the Structure And the Devices Called Vascular Bundles of Plants), E. C. JEFFREY (University of Toronto, later Harvard University), who defined a number of new terms in 1898 like protostele and siphonostele (we will meet them again a little later), as well as the Englishmen G. BREBNER (1902) and F. O. BOWER.
The different ways, in which the terms were used and the independent nomenclature of those times created more confusion than clarity. The situation stabilized around 1910 and an agreement about definitions that went down in the annals as the 'British System' was reached.
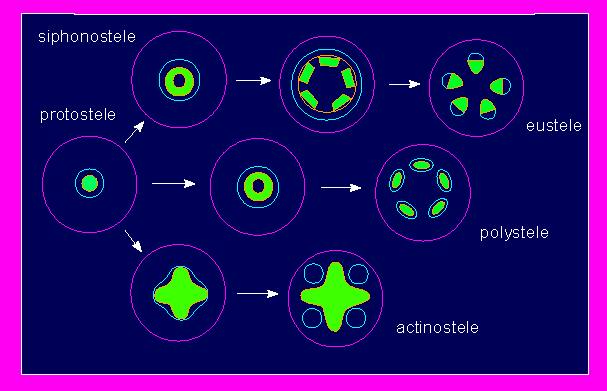
The term stele sums up the vascular system, associated tissues and the pith. It stems, like so many other terms of plant anatomy, from the Greek and means pillar. The theory of steles says that the primary plant body of roots and shoots is constructed according to the same principle, since both contain a central pillar (stele) that is embedded into primary bark. The theory of steles does not deal with secondary growth. The pattern of steles can be regarded as a conservative feature, whose complexity increased in the course of evolution.
The protostele is a conductive system of simple organization. It is a simple, unbranched, centrally located axial strand of xylem coated or interspersed with phloem. A protostele contains no pith. It can be found in primitive vascular plants, like the fossil Rhynia, as well as in young fern shoots, in the shoots of simple aquatic plants and in the roots of most angiosperms. A variation, the plectostele is typical for Lycopodiae.
The siphonostele represents the prototype of the vascular systems of ferns and all further developed vascular (seed) plants. It is composed of several axial vascular bundles that are arranged within the stem in the shape of a tube with enclosed pith.
Leaf and branch traces branch from the axial bundles in direction of leaves and branches and extend into these organs. Leaf traces start always at the point of branching and stretch out to the basis of the petiole. A leaf can be supplied by one or more leaf traces. The axial bundles run along the whole length of a stem section. If you recall how vascular bundles develop during ontogenesis you will understand immediately that they consist of segments that were originally laid out as leaf traces.
The transition from leaf trace to vascular bundle happens as soon as an apically laid out leaf trace gets in contact with the existing vascular system.
Axial bundles are usually interconnected thus creating a communicating conductive system (K. J. DORMER, 1954). The areas, where the leaf traces branch, are the nodes, the in-between sections are called internodes. The structure of the nodes in dicot stems is rather simple, complex linking occurs chiefly in monocots.
Some examples of the organization of vascular bundles will follow. Simple conditions are found in primitive, fossil gymnosperm predecessors (progymnosperms), like in the species Callixylon brownii, whose vascular system was reconstructed by C. B. BECK in 1979. It reveals mostly parallel bundles with only few linkings (anastomoses). A considerable increase in the complexity of the branching pattern occurs in gymnosperms. It is distinguished between open and closed vascular systems, the latter are the leaf traces interconnected by anastomoses. Each leaf of Ginkgo biloba is supplied by two leaf traces that stem from different, but neighbouring axial bundles. The Cycadales like Dioon spinulosum have five leaf traces per leaf that spring from distant axial strands. The leaf traces enclose the cylinder of vascular bundles like a ring, before they branch together in the direction of the leaf (A. DORETY,1919). Here, the organization of the nodes is already rather complex.
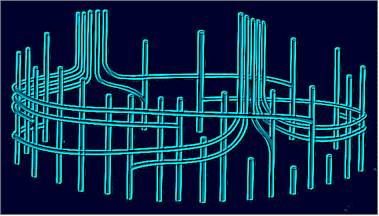
Angiosperms stand out due to the high variability in the construction of their vascular cylinders, although the basic patterns are mostly identical. Nevertheless a serious difference between mono- and dicots does exist. The situation in dicots is closer to the basic pattern and is therefore discussed first.
Variations of the vascular cylinder are based upon direction and height of the helix, upon the number of traces per leaf and the nature of leaf insertion. The term leaf gap that has to be mentioned in this context denotes a gap in the vascular cylinder above the leaf insertion. Angiosperms do as a rule contain five strands of vascular bundles. This number is regarded as a basic unit, of which variations with more or less chords are derived. 91 percent of all species have open systems, closed ones, where several leaf traces and leaf trace complexes are connected are typical for the others (both herbs and wood plants). Here, the basic pattern shows four vascular bundles per vascular chord. Often the leaf gaps are very big, like those of many angiosperms (cacti, Cycadeae, ferns). Their vascular cylinder looks net-like.
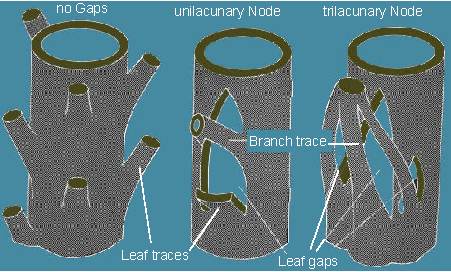
Stele types. On the left: siphonostele without leaf gaps. On the middle and the right: siphonostele with leaf gaps (according to Y. OGURA, 1938).
In summary, the following trends can be determined in the evolution
of seed plants (C. B. BECK et al., 1982):
Monocots provide a highly specialized exception. The vascular bundles of monocots seem to be scattered across the whole stem, being slightly more concentrated at the periphery. If the course of a single axial bundle is examined, a clear helical arrangement can be detected. Never is a vascular bundle exclusively central or peripheral. Depending on where the stem is cut, it can sometimes be found in the centre and sometimes in the periphery. At regular distances leaf traces branch. They, too, run for a long time along the stem, though normally in the periphery. This is also the reason, why particularly many vascular bundles can be seen in the periphery of cross-sections. Part of them are axial chords, part leaf traces. They can usually not be distinguished in cross-section, although axial chords have a slightly greater diameter. Axial bundles may be interconnected by bridging bundles.
In view of this building plan, the statement that the vascular bundles are dispersed over the whole stem, gained by the examination of cross-sections seems to be misleading. Here as well as in dicots, the construction plan shows a clear rotation symmetry with a modified, in cross-section seemingly abolished shape of the vascular bundle. Each vascular bundle is laid out during ontogenesis in a peripheral position and it is not before the following growth that they in parts come to run in the centre.
|
|