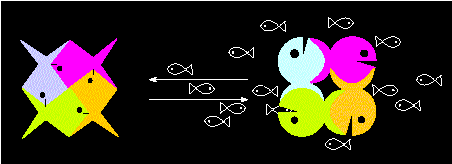The enzyme kinetics reviewed in the two sections above are valid for enzymes of simple structures, i.e. for such enzymes that consist of just one polypeptide chain which again has only one substrate-binding site. In this context, the participation of coenzymes can be neglected. The inhibition kinetics indicate that the molecular conformation of an enzyme is not rigid but has a limited flexibility. Such a reversible deformation occurs as soon as a molecule has bound to the enzyme. It does not matter where it binds. Molecules that induce structural changes of the enzyme are collectively called effectors. Among them may also be substrates.
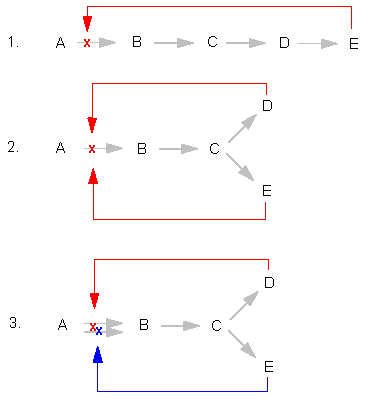
Many enzymes that hold central switch positions in branched biosynthesis pathways can temporarily be shut down by the final product of one or both pathways. This is termed end product inhibition. It assures that the synthesis of a product is stopped as soon as enough has been generated.
|
|
|
|
|
4. Inhibition at a ramification of a biosynthesis pathway (sequential inhibition) |
A rather large group of enzymes consists of several polypeptide
chains. The complex (quaternary structure) may either be formed by
identical or different chains. In such a complex enzyme, the
transmission of information functions far better than between free
polypeptide chains.
The binding of a substrate molecule to, let us assume, one of four polypeptide chains distorts at first this chain's structure. But the distortion is immediately transmitted to the other three chains so that their substrate binding sites are also changed.
"Allosteric model" of a tetrameric protein. The transition between one state of activity to another (transition between two alternative conformations) is elicited by the substrate (the little fishes) and occurs in a co-operative manner (according to M. EIGEN and R. WINKLER, 1975)
Two possibilities exist: either a higher reactivity is achieved (positive co-operation) or a reduced one (negative co-operation, desensibilization). Which alternative applies for a special case can be recognized by its turn-over kinetics. Enzymes of several polypeptide chains that are controlled in this way are called allosteric enzymes.
A positive co-operation generates a sigmoid curve expressing that the fist substrate molecule has been bound with a weak affinity while the following ones are bound by the other chains that are by now in a state of increased reactivity. The kM value of allosteric enzymes is consequently no constant, but a function of the substrate concentration.
A negative co-operation generates a signal that reduces the substrate affinity.
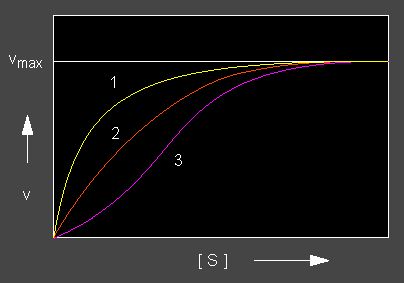
Enzyme kinetics: Yellow curve: Negative co-operation, red curve: nNon-co-operative, lilac curve: Positive co-operation. Enzymes with co-operative effects are always built from several subunits (allosteric proteins).
The ability of enzymes to be controlled is of no lesser importance in
metabolism than their catalytic activity. It decides which of two
alternative pathways is chosen in a certain situation.
Since these control processes are reversible, it is guaranteed that a new decision can be made at each moment so that the cell can react without loss of time to changes in the substrate or product concentration. Viewed from a point of scarce resources and permanent energy crisis, it does become clear that these are mechanisms working very efficiently.
The fast working, reversible property of enzymes to be regulated is based on the instability of weak interactions, since everything said about bonds and the transmission of information elicited by them is caused by their generation and breaking.
Beside this, enzymes may have the property to be regulated by changes of their geometrical structure brought about by the generation or breaking of covalent bonds. Such modifications are without exception irreversible. Among them are the phosphorylation and acetylation of enzymes as well as the breaking of a polypeptide chain that renders an inactive enzyme active. A regulation can also be achieved by the number of enzyme molecules per cell.
|
|