It has been tried for a long time to cultivate plant tissue on artificial nutrient medium. The first approaches go back to the Austrian plant anatomist G. HABERLANDT (1854-1945, professor at Graz, later at Berlin). At first posed the composition of a suitable nutrient medium large problems. It was the Dutch plant physiologist J. v. OVERBEEK who discovered in 1941 that the addition of coconut milk causes a drastic increase in the growth of plant embryos and tissue cultures. Coconut milk is an endosperm product that has under natural conditions, too, a growth-stimulating effect on the developing coconut embryo. The question which components cause the growth stimulation arose immediately.
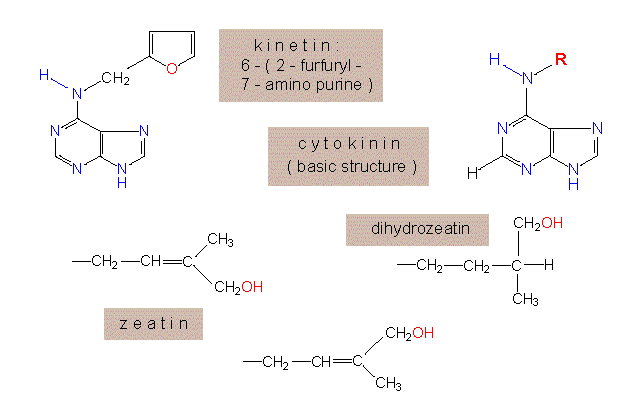
In contrast to auxin is not elongation but growth by cell division stimulated. In 1955 discovered C. O. MILLER and F. SKOOG from the University of Wisconsin at Madison that aged or autoclaved DNA preparations have the same effect, while fresh DNA preparations display no effect at all. In the end was the adenine derivative 6-furfurylaminopurin (= kinetin) identified as the effective substance.
Kinetin is physiologically extraordinary active, although it could not be isolated from any plant cell. Instead was a wide range of similar compounds found. The first that was isolated from a natural source (unripe corn seed) was zeatin (D. S. LETHAM et al., 1964).
Cytokinins is a collective name for compounds of this type. The formulas of these compounds let it appear probable that cytokinins effect the nucleic acids’ metabolism, though this assumption should not be accepted too uncritically. cAMP is known to be a second messenger in animal cells. Although it is derived from ATP and belongs itself to the nucleotides has it only little to do with the regulation of the nucleic acid metabolism in animals (and micro-organisms). Instead have numerous proteins that interact with cAMP been identified. Their conformation and activity changes drastically after binding to cAMP.
In the meantime have several hundred different cytokinin derivatives been synthesized in different laboratories. Many of them are as effective as kinetin. For a biological effect is the substitution of the N-atom no 6 (of ring 6) required. Each exchange of a ring atom causes a drop in activity. An alkyl group substituting N6 has the greatest effect when consisting of a chain of 5 C atoms. A double bond and /or a hydroxy group increase the activity several times. It seems as if all modifications leading to more plane structures would increase the activity. This would explain, too, why the furfuryl ring of kinetin displays such an extraordinary effect. Among the naturally occurring, very active cytokinins is the IPA (isopentenyladenine) from tissue cultures of tobacco. Cells produce it by adding the isopentyl side chain to an adenine residue that is incorporated into tRNA (tRNAser and tRNAtyr).
Cytokinins can – just like auxins – be glycosylated or bound to an amino acid or a protein, thereby being at least temporarily inactive.
Biological activities. Tissue cultures like that of tobacco or maple (Acer pseudoplatanus) develop only after the addition of cytokinin. Beside the rate of DNA replication increase cytokinins, too, the general rate of RNA and protein synthesis. They reduce senescence and stimulate the dark-germination of light-dependent seeds. In addition were several more selective effects observed. Cytokinins
| induce isocitrate-lyase and protease activities in cut pumpkin cotyledons, | |
| induce thiamine synthesis in growing callus cultures of tobacco, thereby removing the thiamine need of the calli, | |
| stimulate auxin synthesis in tissue cultures of tobacco | |
| increase the carboxydismutase and the NADP-glycerinaldehyde phosphatedehydrogenase activities in etiolated rice seedlings | |
| stimulate the development of buds as well as the germination of several seeds, and the accumulation of nitrate reductases in several embryos. |
Cytokinins are usually produced in roots, young fruits, and in seeds. They enter the shoot organs via the xylem. Organs that are cut off from a continuous cytokinin supply like cut shoots age faster than those that are connected to their roots. The addition of kinetin can stop senescence. The development of adventitious roots – and thus the new supply with cytokinins – restores the old state.
The combined effects of IES and kinetin and their relative amounts decide whether a callus tissue of tobacco, for example, differentiates into a root or a shoot. Obviously has an undifferentiated cell two possible routes of development: it can either enter the cycle of enlargement, division, enlargement or it begins to elongate without dividing. A cell that divides often stays mostly undifferentiated, while elongated cells develop a tendency towards differentiation and thus towards specialization. We know that IES alone stimulates elongation while kinetin alone has no effect. Both hormones together stimulate fast cell divisions.
At a ratio of 3 mg/l towards 0.2 mg/l grows a callus tissue (cell divisions take place). If the kinetin amount is lowered to 0.02 mg/l, root development is induced. In case that more kinetin than IES is present (0.03 mg/1.0 mg/l) develop shoots.
The IES/kinetin ratio regulates, among others, also the lignin/pectin ratio in cell cultures of tobacco. At high amounts of kinetin and low amounts of IES is far more lignin than pectin produced, in the opposite case is the lignin/pectin ratio reversed, too.
The interaction of IES and cytokinins is dependent on calcium. Upon addition of calcium is the growth ratio cell elongation/ cell division changed. High concentrations of calcium inhibit the elongation of the cell wall, no new structural elements are deposited.
Just like in the case of auxins have cytokinins, too, no singular mode of action. The variety of hormonally induced phenomenons points to the existence of different receptors.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de