In 1926 studied the Japanese E. KUROSAWA a rice disease that is known as the �foolish seedling�-disease in Japan. The plants grow extremely fast, look spindly and pale and break off easily. KUROSAWA detected that the reason for this abnormal growth is a substance that is secreted by a parasitic fungi (Fusarium moniliforme = Gibberella fujikuroi). It was termed gibberellin.
During the thirties was gibberellin isolated and crystallized by Japanese scientists from Tokyo (YABUTA and SUMIKI ), though it was almost forgotten in the following years. In 1956 isolated C. A. WEST and B. O. PHINNEY a gibberellin from Phaseolus vulgaris and other plants, thus showing that these compounds are far-spread in the plant kingdom. Today are more than 110 different gibberellins known (GA1, GA2,....GA3, GA4.....GA110) that differ only little chemically but very much in their biological activities.
Roughly 30 percent of all known gibberellins are biologically active. All higher plants contain presumably at least one, but usually several active and inactive gibberellins that exist in different concentrations depending on the respective tissue.
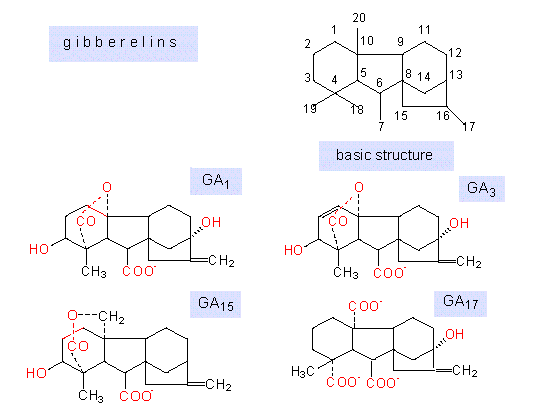
Gibberellins are diterpenoids derived from four isoprenoid units forming a system of four rings. It is distinguished between gibberellins of 19 and such of 20 C-atoms. The twentieth C-atom is not part of the four rings but belongs to a side chain (CH3 in GA12, CH2OH in GA15, CHO in GA19 or COOH in GA28). Both enumeration and the illustration below show, how the single structures differ.
Young, growing meristematic cauline tissue, apical root cells, young fruits, as well as unripe or germinating seeds are all rich in gibberellins. In sun flowers is the gibberellin content highest in the young leaves and in the uppermost internodes while it decreases continuously towards the basal leaves and internodes.
Gibberellin content and growth velocity of a tissue are correlated. Despite the shoot are high concentrations measured, too, in the root tips. The idea that they are produced in the roots seemed logical though indications exist that they are only converted in the roots, i.e. that one gibberellin is transported from the shoot into the root where it is converted another one. The new product is then transported back via the xylem into the shoot. Both in xylem- and phloem exuded matter have gibberellins been detected showing how the distribution mechanism for hormones works in this type of plant. Moreover exist reliable indications that symplastic transport exists in the tissues themselves.
Biological activities.
In a number of plant species is flower formation controlled by extern factors, especially by light (short-day plants and long-day plants) or low temperatures (vernalization). If certain long-day plants or plants requiring vernalization (Hyoscyamus, Daucus, Crepis, Silene) are cultivated under short-day conditions and/or without a cold stimulus, flower formation does not take place. After addition of GA3, though, do they flower even without the otherwise necessary extern signals. This does not mean that gibberellins take actively part in flower formation. It seems more as if flower formation in the mentioned species were dependent on a previous elongation of the stem axis, and that gibberellins stimulate mainly this step. In most other long-day plants (whose stem axis does not elongate immediately before flower formation) and in all short-day plants display gibberellins no effect on flower formation. In plants germinating only in light was it shown that gibberellins are also able to substitute PFR. Gibberellins (like the best-studied GA3) control the formation and secretion of hydrolases in grains (like that of barley): the mobilization of endosperm storage compounds during germination supplies the embryo with nutriments. A number of hydrolases, among them the alpha-amylase, a protease, and a ribonuclease participate in this process. They are produced in the cells of the aleuron layer during the early stages of embryo germination . The way in which this reaction works was elucidated in the laboratory of J. E. VARNER at the Washington University in St. Louis at the beginning of the seventies.
Under the influence of GA3 rises the concentration of active alpha-amylase as a consequence of enhanced alpha-amylase production. Alpha-amylase is uncomplicated to detect and is therefore a suitable marker for the hormone-controlled enzyme production of this system. The production is inhibited by transcription inhibitors (like actinomycin D or chlororamphenicol). Simultaneous to the enzyme synthesis increases the endoplasmatic reticulum in size and new polysomes are produced. Involved is an increased incorporation of phospholipids in membranes as well as the synthesis of enzymes required for the formation of new membranes
The results indicate that GA3 stimulates at first the production of a protein synthesis machinery that produces in a second step the mentioned hydrolases. Here, too, has GA3 a selective effect since an enforced production of alpha-amylase mRNA was detected after the application of GA3.
It should be mentioned though that the activities observed in barley seedlings could be reproduced only with few species though the differentiation steps were the same. It has therefore been asked whether the change in the concentration of GA3 is not just a side effect of the normal process of differentiation (P. HALMER; 1985).
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de