
Just like the comparative analysis of morphological features, chromosomal analysis of numerous species and genera points at phylogenetic relations between single species. Often, the genomes of related species are so similar or almost completely homologous, that even fertile diploid primary hybrids develop:
ABC x A'B'C' > A'BC, AB'C, ABC', A'B'C, AB'C', A'BC'
In numerous further cases, the development of new species can be traced back to the addition of the two parental genomes (allopolyploidy). On the other hand, experiments performed in the 19th century by KÖHLREUTER, GÄRTNER, NAUDIN, MENDEL and others showed, that the hybridization of two species does not necessarily lead to new stable species. Back-crossing of the hybrids with one or both parental species is often possible. As a consequence, parts of the genome of one species can be transferred to the genome of the other species, can enlarge the gene pool and increase the intraspecific genetic variability. Such an interspecific genetic migration is called introgression. Biological species are usually well adapted to the environmental factors of their habitat, and the hybrid is rarely better-suited than the parent. Often, a weakness of the hybrids and a reduction of fertility that is not to be underestimated can be noticed, so that the hybrids are inferior to the original species. They may, nevertheless, be of advantage in the colonization of new, often ‘disturbed’ habitats:
The two species Geum rivale and Geum urbanum are sympatric in Europe. Geum urbanum living in moist forests, hedges, and manmade biotopes flowers yellow, its calyx is folded back. In contrast, Geum rivale occurs in flood-plain and swamp forests and on humid meadows. Its flowers are nutant, the petals are reddish on the outside, the calyx is not folded back. Geum rivale flowers three to four weeks earlier than Geum urbanum. It is a typical bee plant, Geum urbanum with its ‘open’ and erect flowers is less specialized, self-fertilization occurs. If both species occur in the same habitat, hybrid swarms are not rare. The F1-hybrids show a continuous spectrum of transitions between the two parental species. Most hybrids are fertile, and a number of combinations develop in the F2-generation. Consequently, the two species should be just one species. It is very likely that Geum rivale developed in geographic isolation in south-east Europe. Geum urbanum as a less specialized species managed to be successful in man-made habitats. Due to these ecological preferences, the parental populations remain rather stable. Their hybrids have no selective advantage. Two African Delphinium-species provide a similar example.
Adaptive radiation: adaptation to new conditions. In Southern California, Salvia apiana and Salvia mellifera are sympatric species with different ecological requirements. Their flowers are adapted to pollination by different insects. Salvia apiana apiana is mainly pollinated by large carpenter bees (Xylocopa), the pollinators of Salvia mellifera are small to medium-sized bees (Osmia, Apis, etc; K.A. and V. GRANT, 1964). Moreover, a seasonal isolation of the two species exists. Hybridizations of the two species occur hardly ever in natural habitats, while they are more common in areas dominated by humans (C. EPLING, 1947).
Already in 1935, K. M. WIEGAND pointed out, that isolation barriers work well in natural habitats, but do often collapse in human environments. He gives railway embankments as an example for a new habitat. Trains operating on long distances proved to be very effective in dispersing plant seeds and other plant parts. These plants were sometimes not adapted to the conditions at the new habitats. The hybridization with endemic, but related, allopatric species provided a solution.
Adaptation to new pollinators and habitats, changes of flower morphology. In California, three species of the genus Penstemon (beard-tongue, family: Scrophulariaceae) differing in flower shape, size and colour occur. Penstemon grinnellii is common in mountain pine forests. Its blue corolla is two-lipped and wide open. Penstemon centrathifolius has tube-shaped, red flowers. The sunny and dry slopes of Southern California are its area of distribution. The third species, Penstemon spectabilis, lives in a relatively new biotope influenced by climatic changes: on brushwood – covered slopes, substitute of formerly existing forests. Its flowers are blue to slightly purple, their shape is an intermediate of P. grinnelii’s wide open and P. centrahifolius’ tube-shaped flowers.
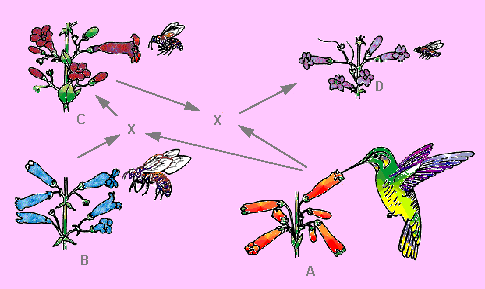
Penstemon grinellii is usually pollinated by large carpenter
bees (Xylocopa), Penstemon centrahifolius by humming birds,
and Penstemon spectabilis by wasps and medium-sized bees.
The coming into being of the last species, Penstemon spectabilis, could be reconstructed as follows. In the past, natural hybrids of the other two Penstemon species had no selective advantage, since the existing habitats were already occupied by the parental species, but when brushwood-covered slopes replaced forests, they found a suitable ecological niche. Wasps and medium-sized bees occurring in these habitats turned into their pollinators. Flowers with their nectar accessible to the pollinators became advantageous. Selection favoured and stabilized now plants adapted to dry environments (in contrast to forests) and flowers adapted to pollination by wasps instead of humming birds.
Different degrees of relationships between species and the ability to produce hybrids. J. CLAUSEN (University of California, Berkeley) analyzed the toleration of the genomes of related, partly allopatric, partly sympatric species of the composite genus Laya during the forties of the last century.
These Laya-species can be assigned to three different groups according to their geographic occurrence. The first group consists of two species, Laya chrysanthemoides and Laya fremantii. L. chrysanthemoides occurs in the San Francisco Bay area, L. fremantii exists more to the east, at the foot of the Sierra Nevada. Three species occurring 300 km south of San Francisco Bay and the Sierra Nevada, Laya jonesii, Laya munzii, and Laya leucopappa, belong to the second group. The third group consists of just one species, Laya platyglossa, indigenous in Southern California. Its area of distribution stretches so far north, that it is sympatric with the species of the first and the second group. It does, nevertheless, colonize other biotopes than the first two species. J. CLAUSEN et al. produced all possible hybrid combinations under controlled conditions and determined the properties and especially the fertility of the progeny. All species can be hybridized with each other. The hybrids are characterized by luxuriant growth. Both within group 1 and within group 2, the fertility of the hybrids is 25 to 30% of the fertility of their parental species. It seems, that the isolation barriers between them are not (yet) stable enough. The fertility of hybrids with one parent from group 1 and the other from group 2 is just 5 – 20 %, i.e. the species of different groups are related less closely than species belonging to the same group. The isolated species of group 3, finally, produces almost without exception sterile hybrids (0.5 – 2 % fertility), i.e. hardly any exchange of genes occurs where it is sympatric with one of the other species.
Morphologic variability and chromosomal variations do not always correlate: Clarkia speciosa occurs in Southern California in numerous varieties differing both in their flower structures and their vegetative organs. The area of distribution of the subspecies Clarkia speciosa nitens is northwards, Clarkia speciosa polyantha adjoins to the south (H. and M. E. LEWIS, 1955). Both species are sympatric in a small geographic section. Hybridization occurs. Independent of classifications due to morphologic criteria, karyotypes of the most different Clarkia speciosa – populations were analyzed. All of them have n = 9 chromosomes. Nevertheless, two chromosomal races differing in seven translocations exist. As a consequence, a ring of 16 chromosomes and a chromosomal bivalent develop during the meiosis of the hybrids. The situation is similar to that of Oenothera, Datura, and Rhoeo (Clarkia belongs just like Oenothera to family of evening primroses, also called Onagraceae). The border between the chromosomal races runs through the area of Clarkia speciosa polyantha. Chromosomal variation and morphological variability of Clarkia speciosa polyantha and Clarkia speciosa nitens are thus independent of each other (W. BLOOM and H. LEWIS, University of California, Los Angeles, 1972).
All given examples show, that interspecific hybrids are only able to become a new species, if several independent conditions stabilizing the new genotypes and isolating them from the original species occur simultaneously or subsequently. A restructuring of the genome as a consequence of translocations, inversions or mutations on one hand and the colonization of free ecological niches on the other hand seem to be very important.
Many closely related species can be sorted into groups of related species, that are able to exchange genes as soon as the barriers effective in nature are loosened or removed, demonstrating, how insecure single barriers of propagation and how flexible plant genomes can be. The percentage of fertile hybrids can be taken as a measure of the relationship of two species or populations:
The Phacelia magellanica – group (area of distribution is North America, families: Hydrophyllaceae related with Solanaceae). Species of several different degrees of ploidy belong to this group. The single species occur as diploid, tetraploid, and hexaploid varieties. A number of the tetraploid species form hybrids relatively easy. The gene exchange between them is therefore more effective than the gene exchange between the varieties with different levels of ploidy belonging to the same species (R. L. HECKARD, University of California, 1960).
Die The genus Madia (endemic in North America, family: Compositae). J. CLAUSEN and his collaborators performed numerous crossing experiments with Madia. Click the last link to see their results. These results explain the different degrees of relationship between the single species. They do also show, what a complex of species with fluid transitions in the degree of hybridization is.
The genus Clarkia. H. and M. LEWIS (1955) studied this genus and their results are well-suited for illustrating the phylogenetic relations between the species of this genus and for arranging the species in a phylogenetic tree. Strictly speaking, it is not exactly a tree, since not only branchings (divergences), but also a high percentage of convergences due to species hybridization occur. Such converging evolution (anastomoses) do usually not happen in a (healthy) phylogenetic tree.
F. EHRENDORFER drew up comparable schemes of the evolutions of the genera of the Dipsacaceae-family as well as for the Achillea millefolium-group.
|
|