 In der Pigmentzusammensetzung der Plastiden (= Chloroplasten,
wegen der Anwesenheit der akzessorischen Pigmente auch Rhodoplasten
genannt) sowie der Anordnung der Thylakoide gibt es mehr Gemeinsamkeiten
zu den rezenten Cyanophyta als zu den Plastiden der übrigen
Pflanzen. Diese Korrelation weist darauf hin, daß die Rotalgenplastiden
anderen Ursprungs als die übrigen Plastiden sind. Die Thylakoide
sind nicht zu mehreren gestapelt, sondern einzeln parallel angeordnet.
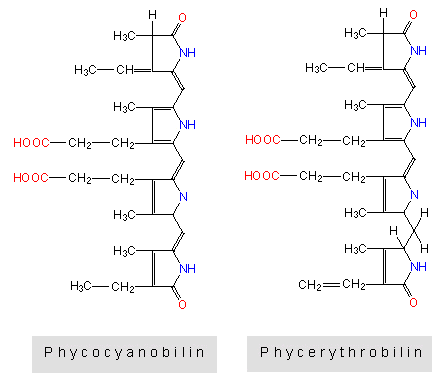
Die Pigmente Phycoerythrin und Phycocyanin sind in besonderen
Aggregaten (Phycobilisomen) gespeichert, die
auf den Thylakoiden liegen .
In der Pigmentzusammensetzung der Plastiden (= Chloroplasten,
wegen der Anwesenheit der akzessorischen Pigmente auch Rhodoplasten
genannt) sowie der Anordnung der Thylakoide gibt es mehr Gemeinsamkeiten
zu den rezenten Cyanophyta als zu den Plastiden der übrigen
Pflanzen. Diese Korrelation weist darauf hin, daß die Rotalgenplastiden
anderen Ursprungs als die übrigen Plastiden sind. Die Thylakoide
sind nicht zu mehreren gestapelt, sondern einzeln parallel angeordnet.
Die Pigmente Phycoerythrin und Phycocyanin sind in besonderen
Aggregaten (Phycobilisomen) gespeichert, die
auf den Thylakoiden liegen .
Begeißelte Zellen wurden nie nachgewiesen. Die meisten der etwa 4000 bis 4500 Arten sind marin, nur etwa 50 bis 100 kommen im Süßwasser vor. In der Regel sind Rotalgen benthisch. Sie wachsen meist auf festen Unterlagen (Felsen, Muschelschalen u.a.) und epiphytisch auf anderen Algen, manchmal auch parasitisch auf nah verwandten, man findet sie daher nur selten an Sandstränden. Die Anheftung an die Unterlage geschieht durch differenzierte, rhizoidartige Zellen.
Das Verbreitungsgebiet umfaßt die Litoralzone aller Weltmeere, in den Tropen sind sie häufiger als in den gemäßigten Zonen, doch kommen sie auch in kalten Gewässern vor.
Rotalgen wurden noch in Tiefen von 100 bis 150 Metern gefunden. Ihre scheinbar eigenartige Pigmentzusammensetzung ist an die Lichtverhältnisse in tiefem Wasser (kurzwellige Strahlung) adaptiert. Eine Anzahl von Arten ist an der Entstehung von Korallenriffen beteiligt. Sie überführen gelöste Calcium- Ionen in unlöslichen Kalk. Ihre Photosyntheseprodukte dienen den Korallen als Nahrung. Fossile, kalkabsondernde Algen gab es bereits im Präkambrium; marine Rotalgen sind seit dem Mesozoikum bekannt. Üblicherweise unterteilt man die Rhodophyceae in zwei Unterklassen
Die Bangiophycidae sind in bezug auf Thallusbau und Fortpflanzungsart die primitivere Gruppe.
Es gibt einzellige und vielzellige Arten mit fädigem oder flächigem Thallus. Zu den bekanntesten Gattungen zählen Bangia und Porphyra. Die Zellen enthalten nur einen sternförmigen Chloroplasten; interkalare Zellteilungen sind vorherrschend. Die Vermehrung erfolgt meist asexuel durch Fragmentation oder durch Monosporen, bei einigen Arten wurde eine sexuelle Fortpflanzung (mit Generationswechsel) beschrieben. Besonderheiten sollen am Beispiel von Porphyra tenera dargestellt werden: Ihr Thallus ist blattförmig, die Kerne haploid. In ihm können sich die männlichen und die weiblichen Gametangien (Spermatangien und Karpogonien (= spezialisierte Oogonien)) entwickeln.
Spermatangien entstehen aus normalen vegetativen Zellen, deren Inhalt durch mehrere aufeinanderfolgende Teilungen in eine Vielzahl von Spermatien aufgeteilt wird. Die Freisetzung einzelner Spermatien erfolgt nach Auflösung der Wand der Ausgangszelle. Karpogone entstehen durch eine äußerlich kaum wahrnehmbare Differenzierung vegetativer Zellen. Freigesetzte Spermatien, die mit den Karpogonen in Kontakt kommen, können mit ihnen zu einer Zygote, einem Karpogon, verschmelzen. Diese teilt sich mehrere Male hintereinander, wodurch ein Karposporangium entsteht, aus dem (diploide) Karposporen freigesetzt werden. Die Karposporen wachsen zu einem fädigen Gebilde aus (Conchocelis-Phase). Da es sich morphologisch ganz erheblich vom Porphyra-Thallus unterscheidet, wurde es ursprünglich als eigene Art beschrieben. Während der Conchocelis-Phase findet die Meiose statt (wann genau?). Dabei entsteht ein Conchosporangium, aus dem haploide Conchosporen freigesetzt werden. Aus ihnen entwickelt sich wieder ein Porphyra-Thallus; der Kreis ist geschlossen. Spermatangien und Karpogonien werden nur unter Kurztagbedingungen (maximal 10 Stunden Licht pro Tag) und bei niederer Temperatur gebildet. Während des Sommers vermehrt sich Porphyra daher rein vegetativ. Die Conchocelis-Phase dient der Überwinterung.
Bangia-Arten verhalten sich in diesem Punkt ganz anders. Deren Conchocelis-Phase ist haploid; die Meiose erfolgt offensichtlich im Anschluß an das Auskeimen der Karposporen. Der Porphyra-Thallus und die Conchocelis-Phase unterscheiden sich nicht nur morphologisch voneinander. Die Zellwände des Porphyra-Thallus sind cellulosefrei, sie enthalten vorwiegend Xylane. Die Zellwand der Conchocelis-Phase hingegen enthält Cellulose (L. S. MUKAI et. al., 1981).
Die Zellwände beider Stadien enthalten Proteine, doch diese Proteine enthalten kein Hydroxyprolin. Auch das ist bemerkenswert, denn ein hydroxyprolinreiches Wandprotein gehört zu den charakteristischen Merkmalen der Zellwände grüner Pflanzen (von Chlamydomonas bis Acer pseudoplatanus (Ahorn)).
Zellen innerhalb des Thallus sind untereinander durch Plasmodesmen verbunden. Bei den Florideophycidae sind jene mit charakteristisch gebauten Tüpfeln versehen. Den Bangiophycidae fehlen sie meist. Eine Ausnahme bilden wieder die Zellen der Conchocelis-Phase.
Florideophycidae sind in jeder Hinsicht höher entwickelt als die Bangiophycidae. Bei vielen Arten sind vor allem die älteren Zellen mehrkernig. Davon ausgenommen sind stets die Geschlechtszellen und ihre Vorstufen sowie die apikal sitzenden teilungsfähigen Zellen. Die Zellwände aller Stadien enthalten Cellulose. Die Plastiden sind meist wandständig und dabei platten- oder bandförmig. Nur Zellen weniger Arten enthalten einen zentral gelegenen sternförmigen Chloro- oder Rhodoplasten.
Das Thalluswachstum erfolgt in der Regel apikal. Das Grundmuster des Thallus besteht aus Fäden, die allerdings zu komplizierten Flechtthalli zusammentreten können, welche durch eine gemeinsame Gallerte zusammengehalten werden. Perikline Teilungen der Abkömmlinge apikaler Scheitelzelle führen zu Verzweigungen, so daß man zwischen Haupt- und Kurztrieben unterscheiden kann. Die Zellen der Kurztriebe sind oft kleiner als die der Haupttriebe, ihre Teilungsfähigkeit ist eingeschränkt. Je nach der Art der Anordnung unterscheidet man zwischen dem einachsigen (uniaxialen) oder Zentralfadentyp, und dem vielachsigen (multiaxialen) Springbrunnentyp.
Beim Zentralfadentyp können eine einreihige Hauptachse und deren Kurztriebe voneinander unterschieden werden, beim Springbrunnentyp besteht der Thallus aus gleichberechtigten, parallel wachsenden Haupttrieben und verzweigten Seitenachsen, die in der Peripherie zu einer gewebeartigen Schicht zusammenschließen, in der die Photosynthese abläuft.
Die Zellen der Fäden sind durch Tüpfel linear miteinander verbunden. Diese sind von einer kontinuierlichen Plasmamembran ausgekleidet und durch einen proteinhaltigen Pfropf verschlossen. Sie erwiesen sich als ein wichtiges Merkmal zur Artunterscheidung innerhalb der Florideophycidae.
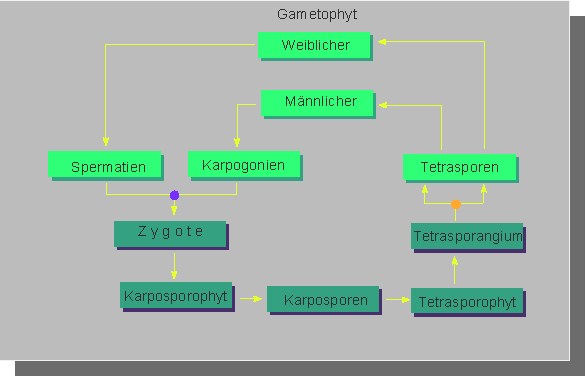
Sexuelle Fortpflanzung ist weit verbreitet. Nach dem Verkleben eines Spermatiums mit der Trichogyne, dem fadenförmigen Empfängnisfortsatz eines Karpogons, und der Einwanderung des Kerns entsteht eine Zygote, die sich zu einem diploiden Karposporophyten (= Gonimoblast) entwickelt Dieser Komplex verzweigter Fäden kann sekundär über Auxiliarzellen ernährungsphysiologisch mit dem Gametophyten in Kontakt treten. Die diploiden und unbegeißelten Karposporen wachsen nach ihrer Freisetzung zu Tetrasporophyten aus, auf denen die Meiose stattfindet. Diese erfolgt in der Regel in Tetrasporangien, die vier haploide Tetrasporen entlassen, aus denen sich wiederum die männlichen und die weiblichen Gamatophyten entwickeln.
Wir haben es bei Rotalgen (übrigens auch bei denen aus der Unterklasse Bangiophycidae) mit einem dreiphasigen Generationswechsel zu tun.
Es gibt
Gametophyt und Tetrasporophyt sind bei vielen Arten gleich gestaltet (isomorph), so z.B. bei den Polysiphonia-Arten, Chondrus crispus u.a. Bei einigen Gattungen (Nemalion, Gloeosiphonia, Gigartina, Asparagopsis und Falkenbergia) kommen hingegen heteromorphe Gameten vor Auch Diözie ist häufig, wobei sich männliche und weibliche Pflanzen unterscheiden können. Ferner gibt es Arten, bei denen die Meiose bereits auf dem Karposporophyten stattfindet ohne daß ein Tetrasporophyt gebildet wird (Liagora tetrasporifera), und es gibt Arten [z.B. die heimische Süßwasserrotalge Batrachospermum (Froschlaichalge)], die einen morphologisch reduzierten "Tetrasporophyten" (einen "Chantransia"- Vorkeim) besitzen, wo die Meiose in einer somatischen (nicht speziell differenzierten) Zelle abläuft, aus der sich der morphologisch dominierende haploide Gametophyt entwickelt. Bilder weiterer Arten / Gattungen: 1. Callithamnion hookeri, und Ceramium deslongchampsii, Ceramium rubrum, Rhodomela subfusca, Cyclostonium purpuraeum, 2. Membranoptera alata und Lomentaria clavelosa, Plocamium cartilagineum, 3. Plumaria elegans
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de