
|
|
|
|
|
HERBACEOUS |
|
Root structure and function
In the mature root the epidermis has lost its root hairs, the cortex is greatly enlarged and serves for food (starch) storage, and the central stele is surrounded by the endodermis. Each cell in this layer is suberized in a (Casparian) strip around the radial walls initially and later over its whole surface. Inside the endodermis are a few layers of cells that remain meristematic, then the phloem and finally the xylem which is often arranged in a band or cross. |
|
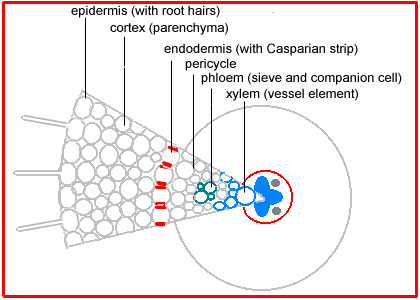 The root apical meristem does not branch. Roots continue their exploration of soil space by developing new apical meristems from the pericycle. The new roots break through the cortex as they grow. |
|
|
|
|
|
Although they are not as diverse as shoots, roots are sometimes modified for special purposes, particularly food storage. Some of our"root crops" are really stems but carrots, parsnips and sweet potatoes are roots. |
|
|
|
|
|
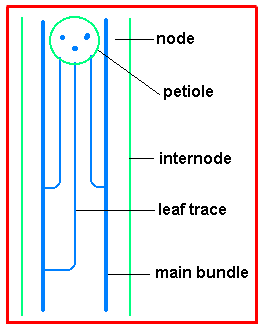
Stem anatomy Leaf primordia arise from the stem apical meristem in a pattern and sequence that determines the arrangement of leaves (singly or in pairs and in one or more planes). In the axil of each leaf primordium is another meristematic area, the bud primordium The node remains a site of meristematic potential, and not just for shoots; roots are also more readily induced to form at nodes than elsewhere. Of course the bulk of stem elongation is internode extension and sometimes this can persist, especially near the base of the internode - intercalary meristems continue stem elongation after other parts have matured. |
|
|
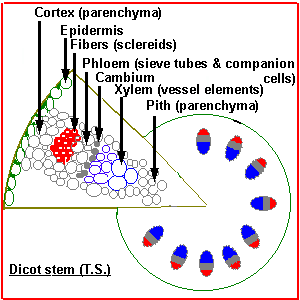
During primary development stems retain their epidermis and usually appear green. Unlike the root epidermis, the stem epidermis contains stomata and there are often hairs or trichomes growing out. Under the epidermis is the cortex of parenchyma cells and within this the vascular tissue. In dicots this can be a cylinder or a ring of discrete bundles. Inside the bundles is pith, a mass of parenchyma cells. In monocots the bundles are scattered through the"ground tissue". |
|
|
|
|
|
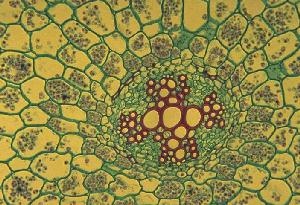
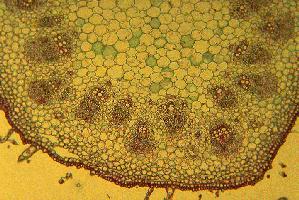
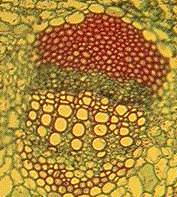
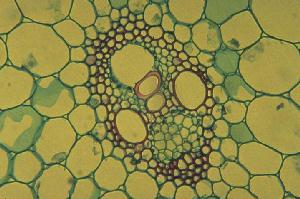
In dicots such as sunflower each bundle consists of phloem on the outside and xylem on the inside of the stem. There are usually some fibers associated with the bundle which can be in a cluster outside the phloem or a sheath around the whole bundle (or both). Across the middle of the bundle is a band of small cells which are capable of cell division. This cambium may extend across the pith rays between the bundles when it is called interfascicular cambium. In monocots such as corn the bundles are scattered, and contain xylem (inside) and phloem (outside) - there are fibers around the outside and the large air-filled cavity in the mature bundle from destruction of the first formed xylem and phloem. There is no cambium and certainly no interfascicular cambium (how could there be?). Monocots have closed bundles because there is no possibility of further differentiation of xylem or phloem. In contrast the vascular bundles of dicots remain open. |
|
|
|
|
|
In primitive plants the vascular system is not so well developed and the lack of connections between different parts of the plant imposes a limitation on water and other movement. During their evolution plants developed ways of linking different structures. At the apex procambium leads to leaf and bud primordia. In a mature dicot stem with discrete bundles, leaf traces branch off from the main bundles below the node and then pass out to the petiole higher up the stem. |
|
|
Underground stems are more common than aerial roots. Many perennials rely on underground stems for their survival through the winter and some of our"root crops", notably white potato are really stems. |
|
|
|
|
|
Copyright © Michael Knee, The Ohio State
University |
|






{kind=link}