
Gefäßpflanzen (Kormophyten und manche Thallophyten) produzieren zeitlebens neue Zellen, und in regelmäßigen Abständen werden neue Organe angelegt. In einem Embryo sind noch alle Zellen gleich teilungsaktiv, doch sobald er eine bestimmte Größe überschritten und ein bestimmtes Differenzierungsstadium erreicht hat, beschränkt sich die Produktion neuer Zellen in der Regel auf bestimmte Bereiche (Meristeme).
Typische Meristeme kommen in Spitzenregionen aller Sprosse und Wurzeln sowie an Haupt- und Seitenachsen vor. Man bezeichnet sie als Scheitel- oder Apikalmeristeme, oder aber als Vegetationspunkte. Die Sproßmeristeme sind meist durch Hüllblätter geschützt und der gesamte Komplex bildet eine Knospe.
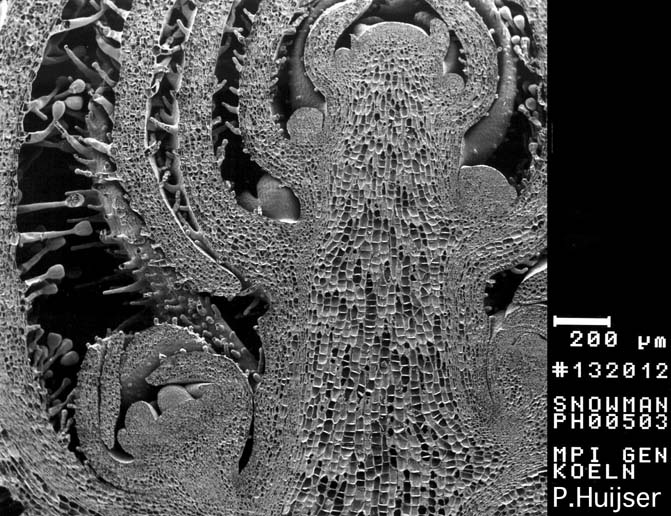
Rasterelektronenmikroskopische Aufnahme: Längsschnitt durch einen Vegetationspunkt (Blütenknospe des Löwenmäulchens (Antirrhinum majus))
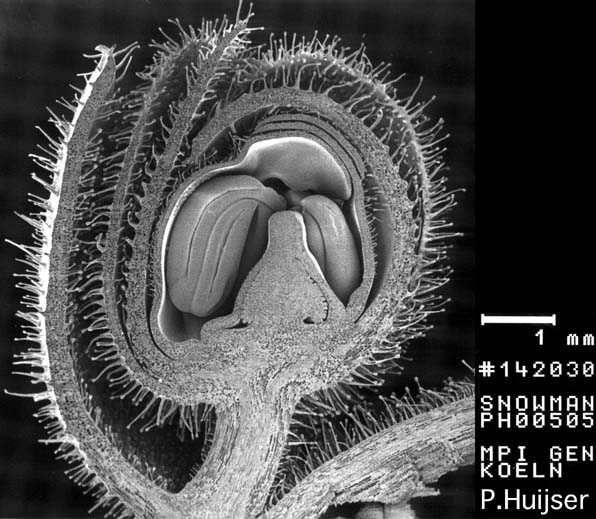
Rasterelektronenmikroskopische Aufnahme: Längsschnitt durch eine reifende Blütenknospe des Löwenmäulchens (Antirrhinum majus)

Aus der Existenz der Apikalmeristeme leitet sich das Spitzenwachstum
als eines der auffallendsten Kennzeichen pflanzlichen Wachstums ab. Dieses
und damit die pflanzliche Entwicklung stehen im krassen Gegensatz zu den
Vorgängen bei der tierischen Entwicklung. Das Wachstum der Tiere ist
nämlich ein allometrischer Prozeß, d.h., es wachsen stets alle
Körperteile gleichermaßen, so daß die Proportionen zueinander
gewahrt bleiben. Sobald ein Tier seine volle Größe erreicht
hat, stagniert das Wachstum. Dennoch finden auch in einem erwachsenen Tier
unentwegt Zellteilungen statt. Man denke dabei nur an die Zellen der Epithelien
oder an blutbildende Zellen. Jedoch wird durch sie lediglich Ersatz für
verbrauchte oder beschädigte Zellen geschaffen. Reparaturen dieser
Art kommen bei Pflanzen nicht vor. Wird z.B. ein Blatt, eine Blüte
oder was auch immer entfernt oder beschädigt, wird das betroffene
Teil weder ersetzt noch repariert. Vielmehr werden an anderer Stelle neue,
oft gleichartige Teile angelegt - und das wiederum kommt bei Tieren nicht
vor.
Abbildung: Vegetationskegel von Elodea
Zellen eines Meristems sind potentiell uneingeschränkt teilungsfähig, was jedoch nicht heißt, daß sie sich ständig in Teilung befinden. Bei vielen Pflanzen kennt man das Prinzip der Apikaldominanz, das besagt, daß Meristeme in Seitensprossen (= subapikal gelegene Meristeme, Seitenknospen) in ihrer Aktivität unterdrückt werden, solange der Hauptsproß wächst. Auch das ist wieder ein Hinweis auf einen Informationsfluß zwischen den einzelnen Meristemen. Wird die Spitze des Hauptsprosses entfernt, entfällt der Hemmeffekt, und die Seitenknospen treiben aus.
Neben den Apikalmeristemen besitzen viele Pflanzen (Gymnospermen, Dikotyledonen) flächig ausgedehnte Lateralmeristeme (Kambium, Korkkambium), die ein sekundäres Dickenwachstum der Sproßachse bewirken. Ihre Aktivitäten können jahreszeitlich schwanken und spiegeln sich beispielsweise in der Ausprägung von Jahresringen wider.
Andere Pflanzen, z.B. viele Monokotyledonen (Gräser u.a.) verfügen über interkalare Meristeme. Das sind aktiv wachsende, vom Apikalmeristem deutlich abgesetzte Meristeme, die zwischen mehr oder weniger differenzierten Geweberegionen, in der Regel an der Basis eines Internodiums, eingefügt sind. Die Bildung sekundärer Meristeme (Folgemeristeme) veranschaulicht, daß Zellen, die einen gewissen Grad an Spezialisierung angenommen und damit die Teilungsaktivität unter normalen Umständen weitgehend eingeschränkt haben, ihren Differenzierungsstatus aufgeben und sich entdifferenzieren. Die Bereitschaft zur Teilung geht also nicht verloren; die Zellen können in den quasi-meristematischen Zustand rücküberführt werden, sobald es die äußeren Umstände erfordern. Die weit verbreitete Anwendung von Stecklingen im Gartenbau oder beispielsweise die Bildung von Sekundärblättern und -wurzeln an Schnittstellen eines Begonienblattes können als Belege hierfür herangezogen werden.

Für uns stellt sich nunmehr die Frage nach der genauen Definition und Abgrenzung eines Meristems, und da beginnen die Schwierigkeiten. Zunächst ein ganz einfaches Beispiel:
Dictyota dichotoma (Gabeltang, eine Braunalge) bildet einen flächigen Thallus aus, dessen Dicke drei Zellschichten umfaßt. Apikal sitzt eine große Scheitelzelle, die sich in der Regel periklin teilt. Die nach unten (subapikal, basalwärts) abgesonderte Tochterzelle teilt sich antiklin, in nachfolgend gebildeten Tochterzellen finden alternierend perikline und antikline Teilungen statt. Der Thallus nimmt damit sowohl an Länge als auch an Breite zu. In regelmäßigen Abständen teilt sich auch die Scheitelzelle antiklin. Als Folge davon entstehen zwei gleichwertige Scheitelzellen, deren Teilungen wiederum zu einer Gabelung des Thallus führen. So einfach das Schema der Teilungen an diesem Beispiel auch erläutert werden kann, so wenig definiert es die Abgrenzung des Meristems gegenüber den ausdifferenzierten Thalluszellen. In der klassischen SACHS'schen Schema (s. Abbildung rechts) eines Kormophyten wird dieser Tatbestand durch einen Gradienten der Teilungsaktivität wiedergegeben. Die meristematischen Eigenschaften gehen nicht von einer Teilung zur nächsten, sondern erst allmählich verloren.
Beispiel 2: Im Gegensatz zu den flächigen Thalli vieler Algen ist der Kormus in der Regel ein dreidimensionaler Körper. Bei einigen Lebermoosen, den Laubmoosen und einfachen Gefäßkryptogamen (Schachtelhalmen und vielen Farnen) bildet eine einzige tetraedrisch gebaute Scheitelzelle (Initialzelle) die Spitze des Vegetationspunkts. Von ihr werden Tochterzellen in regelmäßigem Wechsel der Teilungsebenen nach drei Richtungen (basalwärts) hin abgegeben. Die Tochterzellen wiederum können antikline und perikline Teilungen durchlaufen
Beispiel 3: Bei den Phanerogamen wird die Spitze durch eine ganze Gruppe von Zellen (Initialzellen) ohne jegliches Anzeichen einer Spezialisierung gebildet. Anstelle der Bezeichnung Vegetationspunkt ist es daher zweckmäßiger, den Begriff Vegetationszone oder Vegetationskegel (Beispiele: Acacia, Oxypolis, Weizenähre) zu verwenden. Es zeigte sich zudem, daß das Apikalmeristem mehrschichtig strukturiert ist, wobei man bei den Angiospermen (und einigen Gymnospermen) zwischen außen liegenden Tunikaschichten und dem zentral gelegenem Corpus unterscheidet (A. SCHMIDT, 1924; F. A. L. CLOWES, 1924; A. FÖRSTER, 1943). In den meristematischen Zellen der Tunika laufen vorwiegend antikline, in den Zellen des Corpus sowohl antikline als auch perikline Teilungen ab.
In einigen Abschnitten des Corpus sind bestimmte Teilungsrichtungen vorherrschend, und damit ist bereits ein erster Schritt in Richtung Gewebedifferenzierung getan. Aus der äußeren Tunikaschicht entwickelt sich normalerweise die Epiderrnis. Die übrigen (inneren) Gewebe der Pflanze stammen von Zellen des Corpus, der Tunika oder von beiden ab.
Diese Schemata sind sicherlich eine brauchbare Richtschnur zum Verständnis der pflanzlichen Entwicklung, denn sie weisen auf die Erscheinung einer Positionsinformation hin, die besagt, daß sich eine Zelle nur dann (in eine bestimmte Richtung) weiterentwickelt, wenn sie sich in der "richtigen" Position innerhalb eines Gewebes befindet. Daraus wiederum folgt, daß Zellen, die aufgrund äußerer Einwirkungen (z.B. Beschädigungen) in eine neue Position geraten sind, neue Aufgaben übernehmen, sich also umdifferenzieren. Zu Beginn des 20. Jahrhunderts stellte G. HABERLANDT (Universitfit Graz, später Universität Berlin) den Satz von der Totipotenz der Pflanzenzellen auf. Dieser besagt, daß sich jede Zelle zu Einer ganzen Pflanze entwickeln könne. Wie aber schon die bislang genannten Beispiele zeigen, gilt der Satz für viele, doch nicht für alle Zelltypen. Wir kommen an anderer Stelle noch einmal hierauf zurück (Protoplasten und Gewebekulturen). Doch aus allem folgt, daß ein Wachstum durch Zellteilung zwar für den meristematischen Zustand typisch, doch nicht auf den Vegetations- oder Wachstumsbereich beschränkt ist. Ein Meristem umfaßt keinesfalls nur die Initialzellen und ihre unmittelbaren Derivate, sondern schließt, wie schon angedeutet, verschieden lange Abschnitte des Sprosses mit ein.
Die Situation in Wurzelmeristemen ist im Prinzip ähnlich, obwohl die anatomischen Verhältnisse anders gestaltet sind. Der Vegetationspunkt des Sprosses ist ja, wie wir gesehen haben, durch Hüllblätter geschützt. In der Wurzel übernimmt die Wurzelhaube diase Aufgabe. Sie wird durch Zellen, die vom Meristem kontinuierlich nach außen abgesondert werden, ständig regeneriert. Diese Zellen unterliegen einer hohen Umsatzrate ("turn over"), weil die am weitesten außen liegenden durch wachstumsbedingten Vorschub der Wurzelspitze und Reibung an Erdpartikeln beschädigt und damit verbraucht werden.
Üblicherweise unterscheidet man zwischen dem primären und dem sekundären Sproßwachstum. Ersteres ist mit Sproßverlängerung, Anlage von Blättern und Ausdifferenzierung der einzelnen Gewebetypen verknüpft, letzteres mit (sekundärem) Dickenwachstum der Sproßachse und Anlage neuer (sekundärer) Leitgewebe (A. de BARY, 1877). Obwohl wir uns mit dem Leitgewebe erst später ausführlich befassen werden, sei schon jetzt vermerkt, daß es stets aus den beiden funktionellen Einheiten Xylem und Phloem besteht. Beide zusammen sind zu Leitbündeln vereint. In Sprossen der Samenpflanzen liegt das Xylem innen (in Richtung Achse), das Phloem außen (in Richtung Peripherie), in Blattstielen und Blattnerven liegt das Xylem oben, das Phloem unten; in Pflanzen mit sekundärem Dickenwachstum (fossile Pteridophyten, fossile und rezente Gymnospermen, etliche (verholzte Dikotyledonen) sind sie durch ein meristematisches Gewebe (Kambium) voneinander getrennt. Zusätzlich verfügen viele Pflanzenarten über ein weiteres Teilungsgewebe, das Phellogen oder Korkkambium, das für die laufende Produktion von sekundärem Abschlußgewebe benötigt wird.
Prokambium. Als Prokambium bezeichnet man ein meristematisches Gewebe, das determiniert ist, sich zu primärem Leitgewebe zu differenzieren. Es entsteht dicht unterhalb des Vegetationspunkts und zwar stets dort, wo neue Blattanlagen (Blattprimordien) erscheinen. Im Bereich der Sproßspitze ist die Entwicklung der Leitgewebe daher eng mit der Bildung neuer Blätter verknüpft. Blätter und die mit ihnen verbundenen Leitgewebe werden in einem Wachstumsvorgang angelegt. Die in die Blätter führenden Leitbündel heißen Blattspuren. Die Zellen des Prokambiums sind in der Regel zu Strängen vereint, stellen eine Verlängerung der Leitbündel in den Vegetationspunkt hinein dar und sichern so den Anschluß neu gebildeter Blätter an das Leitgewebe. Die Prokambiumzellen sind gestreckt, sie vergrößern (verlängern) sich im Verlauf ihrer Entwicklung. Das Volumen ihrer Vakuole nimmt beträchtlich zu, so daß sie transparenter (heller) als ihre Nachbarzellen erscheinen. Dieses Stadium wird üblicherweise als der entscheidende Schritt auf dem Weg zur Differenzierung in Leitgewebezellen gesehen.
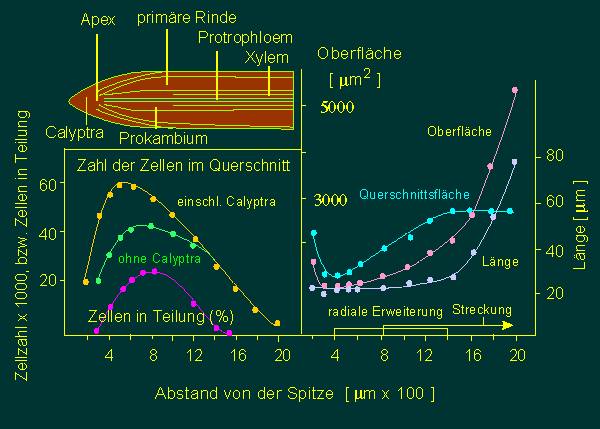
Wachstumsvorgänge in den ersten zwei Millimetern der Wurzelspitze von Allium cepa (Küchenzwiebel). Folgende Bestimmungen wurden durchgeführt, um das Wachstum in den bezeichneten Niveaus zu charakterisieren. Links: Anzahl der Zellen in einem 100 µm dicken Querschnitt,. Rechts: Querschnittsfläche; Länge und Oberfläche der Zellen in verschiedenen Abständen von der Wurzelspitze (Nach W. A. JENSEN und M. ASHTON, 1960).
In Pflanzen mit sekundärem Dickenwachstum verbleiben einige der Prokambiumzellen in meristematischem Zustand und werden damit zu Kambiumzellen. Die bislang knapp skizzierten Merkmale des Prokambiums veranschaulichen, daß es nur einen Übergangszustand repräsentiert. In verschiedenen Höhen (Ebenen) der Sproßspitze kann es daher in unterschiedlichen Entwicklungsstadien vorliegen. Nunmehr stellt sich die Frage nach den Ursachen seiner Entstehung. Zwei Möglichkeiten wären denkbar:
Die Ursache für die Bildung des Prokambiurns ist in Zell-Zell-Wechselwirkungen innerhalb des Gewebeverbandes des Vegetationspunkts zu suchen, welche das Blattstellungsmuster und damit die Position der Leitbündelanlagen determinieren.
Die Induktion geht von ausdifferenzierten Geweben aus. Die Information darüber gelangt über bereits fertiggestellte Prokambiumzellen zum Vegetationspunkt.
Experimentell konnte die erste Annahme bestätigt werden, denn die Bildung und Fortentwicklung des Prokambiums und des Blattstellungsmusters bleiben in isolierten Vegetationspunkten ungestört erhalten. Die Determination der Zellen erfolgt zu einem Zeitpunkt, zu dem noch keinerlei morphologische Veränderungen sichtbar sind. Im Gegensatz zum Begriff Differenzierung, mit dem man eine (meist irreversible) Veränderung von Strukturen und Funktionen in bestimmte spezialisierte Richtungen hin beschreibt, versteht man unter Determination die (ebenfalls meist irreversible) Auslösung dieser Prozesse.
Lange bevor eine sicht- oder meßbare Veränderung erkennbar ist, laufen auf den Ebenen der Genexpression, des Stoffwechsels und der Regulation von Zellaktivitäten Prozesse ab, als deren späte Folge die sichtbaren Veränderungen in Erscheinung treten.
Kambium. Das Kambium ist der Prototyp eines Lateralmeristems. Es ist je nach Herkunft ein- bis mehrschichtig und bildet in der Regel eine zusammenhängende Zellschicht aus, die nahe der Peripherie des Sprosses (und der Wurzel) liegt und die Form eines Hohlzylinders annimmt.
Wo vorhanden, trennt es das Xylem vom Phloem, und wie schon erwähnt, entsteht es im Bereich der Leitbündel aus dem Prokambium, wodurch eine Kontinuität des meristematischen Zustands gewährleistet ist. In den Abschnitten zwischen den Leitbündeln wird das Kambium aus parenchymatischen (also bereits differenzierten) Zellen gebildet. Um dem Rechnung zu tragen, unterscheidet man zwischen dem faszikulären und dem interfaszikulären Kambium (= Kambium innerhalb von Leitbündeln und Kambium zwischen den Leitbündeln). Das interfaszikuläre Kambium muß demnach als ein Folgemeristem oder als ein sekundares Meristem angesehen werden. Einige wenige Monokotyledonen (Drachenbaum [Dracaena], Yucca, Aloe u.a.) besitzen ein mehrschichtiges extrafaszikuläres Kambium.
Die Zellen des Kambiums bezeichnet man oft auch als Initialen, weil sie nach Teilung die Bildung spezialisierter Zellen (aus jeweils einer der Tochterzellen) initiieren. In der Regel kommen im Kambium zwei Zelltypen vor:
Fusiforme Initialen: Das sind plattenförmige, langgestreckte, an den Enden spitz auslaufende, stark vakuolisierte Zellen, die wegen ihres spindelförmigen Aussehens fusiform genannt werden. Aus ihnen gehen die Elemente des Xylems und Phloems, sowie alle übrigen, parallel zur Organachse angeordneten Zellen hervor.
Strahlinitialen: Das sind nahezu isodiametrische, kleine Zellen, die oft zu Gruppen vereint sind. Sie entstehen aus fusiformen Initialen oder deren Teilungsprodukten. Aus ihnen entwikkeln sich in Holzpflanzen radiär angeordnete Markstrahlen (transversale Elemente).
I. W. BAILEY, der in den zwanziger Jahren die grundlegenden Arbeiten über den Aufbau des Kambiums durchführte, konnte bei einem Vergleich eines einjährigen und eines sechzigjährigen Stammes von Pinus strobus (Weymouthskiefer) eine Anzahl relevanter Daten ermitteln. Diesen Werten ist zu entnehmen, daß sowohl die Länge der fusiformen Initialen als auch ihre Zahl mit steigendem Alter des Stammes zunehmen. Die Zunahme der Zellzahl und darnit die Weitung (Dilatation) des Kambiumhohlzylinders beruht auf der durch das Kambium selbst verursachten Verdickung des zentral gelegenen Xylemzylinders.
Durch tangentiale (perikline) Teilungen fusiformer Initialen werden sekundäre Leitelemente in entgegengesetzte Richtungen abgegeben, wobei Xylemelemente nach innen, Phloemelemente nach außen hin angelegt werden. Die Derivate der Initialen sind in radiären Reihen angeordnet, so daß ihre Abstammung leicht zurückzuverfolgen ist. Durch regelmäßiges Auftreten antikliner Teilungen der Initialen werden Defizite, die durch Zunahme des Stammumfangs bedingt sind, ausgeglichen. Die Kambiumtätigkeit ist von jahreszeitlichen Temperaturschwankungen abhängig, was zur Ausbildung von Jahresringen führt. Unterschiede in der Dicke der einzelnen Jahresringe belegen, daß das Ausmaß des sekundären Dickenwachstums durch äußere Faktoren, wie Temperatur, Länge der Lichtphase, Erdfeuchtigkeit u.a. beeinflußt wird. Neben der Produktion von Xylem- und Phloemelementen sowie von parenchymatischen Zellen (in interfaszikulären Abschnitten) fällt dem Kambium eine wichtige Funktion bei der Wundheilung zu.
Korkkambium oder Phellogen. Hierbei handelt es sich um ein sekundäres Lateralmeristem, das der Bildung eines sekundären Abschlußgewebes (Borke; Phellem oder Kork) dient. Es wird ausnahmslos nach außen abgegeben. Vielfach, doch nicht immer, gibt das Phellogen auch Zellen ins Stamminnere ab, die ebenfalls eine Schicht (Phelloderm) ausbilden.
Das Phellogen ist im Vergleich zum Kambium recht einfach strukturiert. Die Zellen erscheinen im Querschnitt rechteckig, in radialer Richtung flach und in tangentialer plattenförmig. Das Plasma ist stark vakuolisiert und es kann Gerbstoffe und Chloroplasten enthalten.
Das Phellogen entsteht aus Zellen der Epidermis und / oder Zellen darunterliegender Parenchymschichten. Man unterscheidet daher auch hier zwischen primärem und sekundärem. Phellogene können demnach im Stamm wiederholte Male angelegt werden. Bei einigen Arten ist es im ersten Jahr einschichtig, später mehrschichtig. Es kann mehrjährig, teils sogar lebenslang, oder auch nur einjährig aktiv sein. Wie beim Kambium wird die Aktivität durch externe Faktoren gesteuert.
|