

Vom Zeitpunkt
der Keimung an zeigen Wurzeln das Bestreben, abwärts, Sprosse das
Bestreben aufwärts, zu wachsen (anisotropes Wachstum). Ein keimendes
Samenkorn kann mehrfach umgedreht werden, doch stets wird sich die Wurzel
nach unten (abwärts) krümmen. Wir haben es daher auch hier wieder
mit einem echten Tropismus, einer Bewegung auf einen auslösenden Reiz
hin zu tun. Daher nun die Frage: Was ist der steuernde Reiz?
"Es ist nicht die Feuchtigkeit des Bodens, die die Richtung der Wurzel bedingt, denn wenn man eine junge Pflanze in eine mit Erde gefüllte Röhre setzt, deren oberer Teil feucht und der untere trocken ist, so steigt dennoch die Wurzel abwärts und der Stengel aufwärts. Setzt man die Pflanze in eine Röhre mit Wasser und erleuchtet den unteren Teil der Röhre, während der obere verdunkelt wird, so ändert sich die Richtung des Wurzelwachstums nicht, ein Beweis, daß sie auch nicht vom Licht abhängt." (A. de CANDOLLE 1834/38)
1806 stellte der britische Physiologe A. KNIGHT durch einen entscheidenden Versuch klar, daß die Richtung des Wurzelwachstums von der Schwerkraft (Gravitation der Erde) gesteuert wird: Er befestigte Keimlinge auf einem senkrecht stehenden Rad, welches er durch ein kleines Mühlrad von einem rasch fließenden Bach um seine horizontale Achse drehen ließ. Durch die Drehung kam die Zentrifugalkraft zur Wirkung. Den gleichen Effekt erzielte er, als er die Keimlinge auf einem sich horizontal drehenden Rad befestigte. Bei schneller Rotation des horizontal gestellten Rades wuchsen alle Wurzeln nach außen (in Richtung der Zentrifugalkraft), bei langsamer Drehung nahmen sie vom Rand an etwa eine Stellung von 45 Grad ein (Resultante im Kräfteparallelogramm: Zentrifugalkraft gegen Gravitation).
KNIGHT meinte zunächst, die Wurzelspitze würde durch ihre eigene Schwere die Wurzel nach unten ziehen, doch konnte diese Annahme schon frühzeitig widerlegt werden, weil die Abwärtsbewegung auch dann auftrat, als das Gewicht der Wurzelspitze durch ein Gegengewicht kompensiert wurde (JOHNSON, 1828).
A. B. FRANCK führte 1868 den Begriff Geotropismus ein, setzte ihn gegen den Phototropismus ab, und unterschied drei Formen:
![]() positiver Geotropismus (wie eben beschrieben),
positiver Geotropismus (wie eben beschrieben),
![]() negativer Geotropismus und
negativer Geotropismus und
![]() transversaler Geotropismus.
transversaler Geotropismus.
Positiver und negativer Geotropismus zusammen werden als Ortho-Geotropismus bezeichnet. Er bedingt die senkrechte (orthotrope) Orientierung der Pflanzenachse. Das Sproßwachstum ist in erster Linie als negativ geotrop zu bezeichnen, denn Sprosse wachsen auch bei völliger Dunkelheit aufwärts. Der Phototropismus ist daher als ein sekundärer, dem negativen Geotropismus meist gleichgerichteter Reiz zu verstehen. Unter transversalem Geotropismus versteht man Wachstumsrichtungen, die senkrecht zur Pflanzenachse stehen. Die Richtung vieler Seitentriebe, Nebenwurzeln, Blätter u.a. wird hierdurch beschrieben; unter plagiotrop faßt man all jene Richtungen zusammen, die von der Orientierung der Sproßachse abweichen; orthotrope Organe (Sproßachse, Hauptwurzel) sind in der Regel radiärsymmetrisch gebaut, plagiotrope (Blätter, Nebenwurzeln u.a.) dorsiventral.
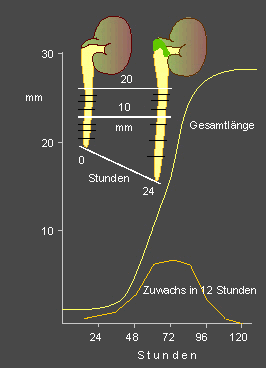 J.
v. SACHS baute die von KNIGHT begonnenen Experimente aus und ergänzte
sie durch die Erfindung des Klinostaten (1879),
mit dessen Hilfe er die Krümmung zum Stehen bringen konnte (Kompensation
der Gravitation durch die Zentrifugalkraft). Durch Kennzeichnung der einzelnen
Wurzelabschnitte mit Tuschestrichen (heutzutage durch andersartige Markierung)
zeigte er, daß sich auch bei der Wurzel nicht die Spitze selbst,
sondern ein darüberliegender Abschnitt (eine Wachstumszone) verlängert.
Pflanzenanatomische Untersuchungen belegten, daß die einseitige Verlängerung
auf einem Streckungswachstum der Zellen beruht.
J.
v. SACHS baute die von KNIGHT begonnenen Experimente aus und ergänzte
sie durch die Erfindung des Klinostaten (1879),
mit dessen Hilfe er die Krümmung zum Stehen bringen konnte (Kompensation
der Gravitation durch die Zentrifugalkraft). Durch Kennzeichnung der einzelnen
Wurzelabschnitte mit Tuschestrichen (heutzutage durch andersartige Markierung)
zeigte er, daß sich auch bei der Wurzel nicht die Spitze selbst,
sondern ein darüberliegender Abschnitt (eine Wachstumszone) verlängert.
Pflanzenanatomische Untersuchungen belegten, daß die einseitige Verlängerung
auf einem Streckungswachstum der Zellen beruht.
Jetzt blieben noch die Fragen zu beantworten, wie der Reiz wahrgenommen und wie er weitergeleitet wird.
1892 postulierte F. NOLL (1858-1908, Prof. in Bonn, später in Halle), daß es in Zellen der Wurzelspitze mikroskopisch kleine, bewegliche Teile geben müsse, die einen Druck auf das Plasma der jeweiligen Unterseite der Zelle ausüben. Der Druckreiz müßte dann vom Plasma in eine Dehnung der Zellwand und in stärkeres Wachstum umgesetzt werden. Eine Bestätigung fand die Annahme (Statolithentheorie) durch die Pflanzenanatomen B. NEMEC in Prag und den Österreicher G. HABERLANDT, die in den Zentralzellen der Kalyptra die beweglichen und wirklich der Schwerkraft folgenden Partikel sahen und sie als Amyloplasten (Stärkekörner) identifizierten.
In einem Punkt bleibt die NOLLsche Annahme angreifbar, denn durch eine Verlagerung von Statolithen kommt es ja nicht zu einer Zellwanddehnung inden stärkehaltigen Zellen, sondern zu einer Streckung von Zellen im Bereich der Wachstumszone. Daraus ergibt sich zwangsläufig die Frage nach der Reizweiterleitung. Eine definitive Antwort steht aus, doch wieder ist Auxin mit im Spiel, wenngleich das Verteilungsmuster keineswegs mit dem beim Phototropismus beobachteten identisch ist. Für seine Mitwirkung sprechen die folgenden Feststellungen:
Von der Spitze zur Basis nimmt die Verlagerungsfähigkeit der Amyloplasten und ebenso der Einfluß der Schwerkraft auf den Auxintransport ab. Durch Anwendung starker Fliehkräfte können auch im basalen Bereich Amyloplastenverlagerung und zugleich Auxinquerverschiebungen festgestellt werden.
Mutanten mit kleineren Amyloplasten zeigen eine geringere Verlagerungsrate derselben im Schwerefeld, geringere Einflüsse der Schwerkraft auf den Auxintransport und geringere geotrope Krümmung. Letztere bedeutet nicht eine geringere Krümmungsfähigkeit der Mutante schlechthin, denn bei der phototropen Reaktion wird kein Unterschied zwischen Mutante und Wildstamm beobachtet.
Bei der Besprechung des Phototropismus wurde das Verhalten der Phycomyces-Sporangien erwähnt. Ein in gewisser Hinsicht vergleichbares Beispiel ist für den Geotropismus (Gravitropismus) bekannt, nämlich die Reaktion der einzelligen Rhizoide der Armleuchteralge (Chara). Sie sind transparent, gegenüber Licht indifferent, und Verschiebungen von Inhaltsstoffen sind mikroskopisch leicht zu verfolgen. Besonders auffallend ist die Existenz stark lichtbrechender Zelleinschlüsse (Glanzkörper) im Bereich der Rhizoidspitze, deren Verlagerung mit einer geotropen Krümmung korrelierbar ist (J. BUDER, 1961). Auf dieser Beobachtung aufbauend, haben A. SIEVERS und Mitarbeiter (Botanisches Institut der Universität Bonn) versucht, durch mikroskopische, elektronenmikroskopische und biochemische Analysen einen unmittelbaren Zusammenhang zwischen der Verlagerung der Glanzkörper und der geotropen Krümmung zu erstellen. Durch Versuche, in denen Mikrofilamentaktivitäten durch Cytochalasin B ausgeschaltet wurden, wurde gezeigt, daß die Statolithenverlagerung ein aktiv gesteuerter Prozeß ist, an dem die Mikrofilamente beteiligt sind. Die elektronenmikroskopischen Aufnahmen ergaben, daß Golgi-Vesikel am Zellwandwachstum mitwirken und daß sie beim Einsetzen der Krümmungsreaktion in diejenigen Zellbereiche verlagert werden, in denen Zellstreckung stattfindet. Zusammenfassend konnten die folgenden Schlüsse gezogen werden:
Die Statolithen hemmen das Zellwandwachstum in denjenigen Zellabschnitten, in denen sie nahe dem Plasma liegen (also genau umgekehrt, wie es NOLL postuliert hatte). An Stellen, auf die Statolithen zuerst sedimentieren, kommt es zu einer leichten Eindellung. |
|
Golgi-Vesikel sammeln sich an der gegenüberliegenden Flanke (Seite). Sie sind an der dort einsetzenden Zellwandsynthese, die zur Krümmung führt, beteiligt.Durch Belastung der Wurzelspitze mit Harzkügelchen läßt sich die Wachstumsrate der einzelnen Abschnitte genau verfolgen. |
|
Wenn Statolithen in einem vertikal wachsenden Rhizoid verlagert werden, wird das gleichmäßige longitudinale Wachstum inhibiert. |
So eindrucksvoll die Befunde an Chara auch sind, so wenig erklären sie die Kausalkette in mehrzelligen Wurzeln; denn dort finden Reizperzeption und gravitrope Reaktion in unterschiedlichen Zellen statt, die darüber hinaus auch noch durch andere, nicht reaktive Zellen voneinander getrennt sind. Die Ergebnisse belegen jedoch eine andere Tatsache: Sie zeigen nämlich, daß die black box zwischen Reizperzeption und Reaktion aus unterschiedlichen Systemelementen aufgebaut sein kann.
|
|