
Storage proteins are a group that comprises proteins generated mainly during seed production and stored in the seed that serve as nitrogen sources for the developing embryo during germination. It is obviously more effective for the plant to use proteins instead of secondary plant products for this purpose. The average protein content of cereal grains is 10-15 % of their dry weight that of leguminose seeds 20-25, while it is only 3-5 % in normal leaves. Besides seeds, storage proteins can also be found in root and shoot tubers, like in potatoes.
No clear definition what a storage protein is, exists. The term is operational and was coined for all those proteins whose share in the total protein amount of the cell is greater than five percent. Usually, the following properties can be found, too:
|
|
The proteins have no enzymatic activities. |
|
|
They serve as nitrogen sources for the germinating seed. |
|
|
They occur normally in an aggregated state within a membrane surrounded vesicle (protein bodies, aleuron grains). |
|
|
They are often built from a number of different polypeptide chains. |
Storage proteins are important for human nutrition (plant proteins) and numerous studies concerning their structure and biosynthesis have therefore been published during the last years. Furthermore, an interest in the production of mutants with an increased protein content or an increased amount of essential amino acids exists.
It will be shown later that genetic engineering enables the analysis of gene structures and gives answers to the following questions:
|
|
how many genes encode a specific protein or |
|
|
what relation exists between the final product and the structure of the corresponding gene (= the respective section of the DNA) ? |
A simplifying rule says that leguminoses contain mostly two types of storage proteins, legumin and vicelin. The legumins - as well as the vicelins - are very similar in the different leguminose species. Gramineae contain a third type: prolamin and, depending on the origin, it is distinguished between the zeines (from Zea mays), the hordeines (from Hordeum vulgare) etc.
In contrast to legumins and vicelins that are mainly located in the cotyledons of seeds, prolamines are found in the endosperm.
Legumin. The first detailed studies on plant storage proteins were carried through by T. B. OSBORNE (Connecticut Agricultural Experimental Station) at the end of the last century, an interim report was published in his book 'The vegetible proteins ' in 1924. OSBORNE could prove that two protein fractions, legumin and vicelin, are present in the seeds of Pisum sativum and that similar proteins can be isolated from the seeds of other leguminoses like Phaseolus vulgaris or Glycine max.
Further studies carried through in the following years showed that legumins occur in the seeds of many dicot families and that a compound similar to legumin is also produced in monocots. These molecules are usually polymers with molecular weights around 300,000 to 400,000. They are typically constructed from two subunits, the acidic and the basic polypeptides. The quaternary structure is composed of six acidic and six basic polypeptides that are linked by disulfide bonds. This rises the question how it is that both polypeptides occur in the plant always in equimolar amounts and how the proteins are brought into the vesicle.
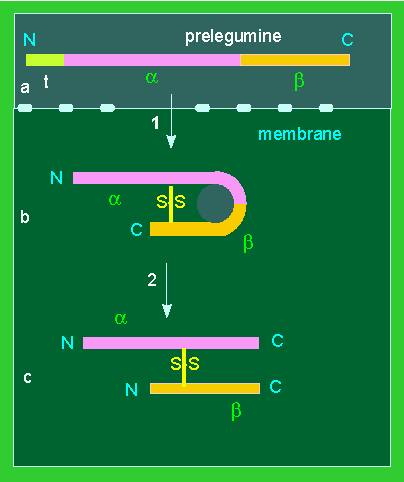
Legumine synthesis: Posttranslational processing. A. Preprolegumin is the complete, primary translation product. The N-terminal transit sequence (t) is needed for the passage through the membrane, it is cleaved off afterwards. B.A disulphide bond is generated. Prolegumin is formed C. The polypeptide chain is cleaved into the two legumin chains alpha and beta. They stay together due to the disulphide bond. A complete legumin complex contains six such units (according to K. MÜNTZ et al., 1985, changed in 1988).
It is known from the studies of secretory proteins of animals that they are produced as precursors. The polypeptide chain has a signal sequence (or transit sequence) of around 20 mostly hydrophobic amino acids at its N-terminus. The polypeptide chain that is at first not folded is synthesized through the membrane. After passing the membrane, the signal sequence is cleaved off proteolytically. This mechanism is also used by plant legumins. Furthermore, the newly synthesized polypeptide chain is proteolytically cleaved into two parts linked by a disulfide bond. One is acidic and one basic. The final molecule is thus the product of a step-by-step processing of the primary gene product. A similar situation is known to exist with insulin. Here, too, the A- and B-chains are cleavage products of an original single chain.
Vicelin. Vicelins are glycoproteins with 1-5% neutral sugar residues. They, too, are constructed from several polypeptide chains that are united by a quaternary structure. All chains are cleavage products of one primary gene product.
Prolamines. Prolamines, the storage proteins of gramineae, are characterized by a high proportion of proline and glutamine. They are soluble in alcohol. They do also have a signal sequence but in contrast to legumines and vicelines, the polypeptide chain is not cleaved. They are the products of a number of around 30-100 genes. Contrary to the genes of the leguminose storage proteins, they do not contain introns, i.e. each gene consists of one undivided translated DNA chain. Among the best-known prolamines are the zeines, the storage proteins of maize. They make up to 50% of the total endosperm protein content. Zeines are polypeptide chains with molecular weights of 10,000, 15,000, 19,000, 22,000 and 27,000. The chains of the 22,000 and 19,000 proteins consist of several (up till eight) serial, similar amino acid sequences:
Each of these domains consists of an alpha-helix. The alpha-helices are connected by non-helical DNA. The tertiary structure of the molecule resembles a heap of parallel cylinders. The homologous domains are the products of gene sequences that originated by multiple duplication of a short original gene and that were subsequently combined in tandem to one gene. Numerous such cases have been described in animals. Zein and WGA are impressive examples for the existence of this mechanism in plants. Zeins are poor in lysine and tryptophane and are thus less valuable for human nutrition than proteins with a higher content of essential amino acids.
As has been mentioned before, maize mutants with higher contents of lysine have been cultivated (opaque-2, opaque-7, floury-2, and others). They have no altered zein structure, but a strongly reduced zein content. The relative amount of other storage proteins with a higher lysine content is thus increased. The total protein content is smaller than in 'normal' maize.
|
|