 ERNST
STAHL (Jena),
ERNST
STAHL (Jena),
one of the pioneer researchers of secondary plant compounds.
Plant cells contain far more compounds than are produced by the basic metabolism. Terpenes, waxes, alkaloids and pigments are just some key words that illustrate what is meant. On the suggestion of A.KÖSSEL (1891), it is distinguished between basic and secondary metabolism. The term basic metabolism comprises all pathways necessary for the survival of the cells, while secondary plant products are such that occur usually only in special, differentiated cells and are not necessary for the cells themselves but may be useful for the plant as a whole. Examples are flower pigments and scents or stabilizing elements. Both metabolisms are overlapping since it is often not understood why a certain compound is produced. Furthermore, no typical cell exists so that some compounds may be necessary for the survival of some cells but not for that of others.
Plant cells produce a vast mount of secondary products. Many of these are highly toxic and are often stored in specific vesicles or in the vacuole. Several studies indicate that this kind of storage functions on one hand as a detoxification of the plant itself and generates on the other hand a reservoir of, for example, nitrogen-rich molecules. In contrast to animals, these are not excreted by plants. Some secondary plant products can be reversibly degraded and are fed into the basic metabolism while others cannot.
Although secondary plant products are very common, this does not mean that every plant can produce every product. Some compounds are restricted to single species, others to related groups. But they are nearly always found only in certain specific plant organs, often in just one type of cell (and there again only in a certain compartment). Also, they are often generated only during a specific developmental period of the plant.
Good reason exists for the use of some secondary compounds (or, even better, groups of chemically similar compounds) as features of classification. But what is true for other morphological features, is also true for these: the presence of a chemical substance in a plant is adaptive although this may not always be as clear as in the case of flower pigments, lignin or cutin.
Many secondary compounds have signalling functions. Among them are plant hormones that will be dealt with elsewhere due to their importance. They influence the activities of other cells, control their metabolic activities and co-ordinate the development of the whole plant. Other substances like the just mentioned flower colours serve to communicate with pollinators or protect the plants from feeding by animals or infections. Some plants, for example, produce specific phytoalexines after fungi infection that inhibit the spreading of the fungi mycelia within the plant. A number of substances is secreted and influences the existence of other species. Especially with algae but with fungi, too, the communication of the cells is stimulated or kept upright by extracellular substances. These substances may be of consistencies ranging from gas to jelly-like. Small excreted molecules have large losses due to diffusion though they are also marked by a wide range of action. Many of them are antibiotic i.e. they inhibit the existence of competing species in the surrounding of their producer thus safeguarding its ecological niche.
The mutual influence of plants by secretion is called allelopathy. It occurs not only in algae but also in higher plants.
Allelopathic substances may damage the germination, growth and development of other plants. Their influence is only rarely stimulating. Insects (and other animals) have developed defence strategies against the insecticide effects of some secondary plant products. During evolution, at first detoxification mechanisms, later even dependencies on certain plant products were developed. Some species, for example, need starting compounds for their steroid synthesis that were originally meant to be a plant defence. They are slightly modified within the animal and get thus a simpler structure.
Since centuries, many plant compounds have an outstanding role in medicine. Their pharmacological - and consequently also their economical - value has lost nothing of its importance until today. They are used either directly or after chemical modification. It should be mentioned that some plant products have psychopharmacological effects and morphine or mescaline are even counted among the 'hard' drugs.
Some secondary compounds are not only found in phylogenetically related species (or families) but also in species that are not directly linked. Even J. v. SACHS mentioned in his 'Lectures on Plant Physiology' ("Vorlesungen über Pflanzenphysiologie", 1882) that a number of species that cannot be phylogenetically related develops latex ducts. Though he noticed, too, that the latex ducts differ both in their morphology and in their ontology. We know today of a range of metabolic compounds that have developed in parallel in non-related species and are produced by different biosynthetic pathways.
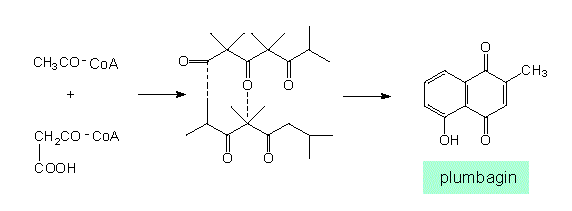
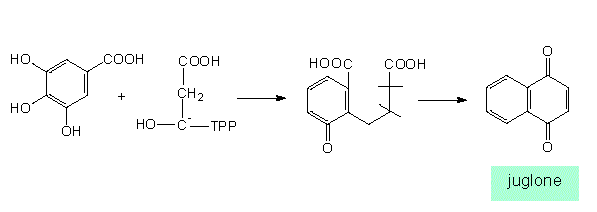
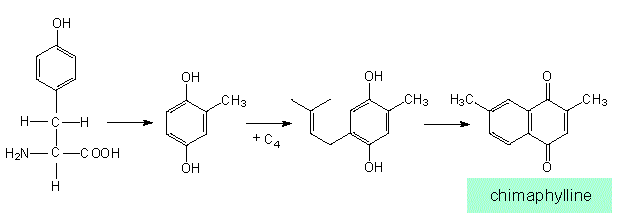
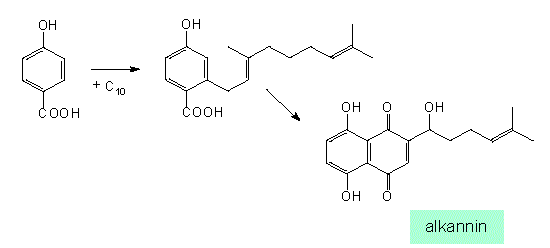
Similar products (in this case naphthoquinone derivatives) can be obtained from very different starting substances and can be synthesized via different intermediates (according to J. B. HARBORNE, 1977):
The acetate-malonate pathway. Synthesis of plumbagin (Plumbago, Plumbaginaceae)
The shikimate pathway, synthesis of juglon (Juglans, Juglandaceae)
The homogentisate pathway. Synthesis of chimaphylline (Chimaphila, Pyrolaceae)
The hydroxybenzoate pathway. Synthesis of alkannin (Plagiobothrys, Boraginaceae)
A number of mutants are unable to synthesize certain products due to a blocked metabolism. A classic example are the white-flowering varieties of species with otherwise coloured, often red flowers. Perhaps, it is indeed a feature of some synthetic pathways of the secondary metabolism that genetic defects are permissible and that their carrier has often a large probability to survive under natural conditions.
The chemical structure of secondary plant products is without exception more complex than that of primary products. This becomes comprehensible when remembering that many though by far not all of them are derived from amino acids or nucleotides. Most of the compounds found in plants belong to rather few families of substances. Only small chemical modifications like methylations, hydroxylations, intercalations with metal ions, etc. lead to a wide spectrum of functionally different substances.
The most important substance families are:
These substances are often glycosylated (certain sugar residues have been added) so that their water solubility is increased. The sugar-free part of such a substance is called an aglucon.
During the last 20 to 30 years, the analysis of secondary plant products has progressed a lot. The use of modern analytical techniques like chromatography (in all its variations), electrophoresis, isotope techniques and enzymology succeeded in the elucidation of exact structural formulas and the most important biosynthetic pathways.
Among the largely unanswered questions are those after the compartment a certain substance is produced in, is that whether all steps of the biosynthetic pathway take place in this compartment or only the first and the last, whether synthesis and storage occur in different compartments, how organ-specific occurrence can be explained and how biosynthesis is controlled. Are the biosynthetic pathways ramified, do they lead to several, related products, exists a feedback inhibition? Why do the pathways of related species differ? How are the synthesis and the activity of the necessary enzymes controlled? Are they monomerous proteins or multienzyme complexes, do they occur in a free state or are they bound to a membrane? How are they transported within the cell and are they possibly excreted?