
A whole range of different membranes exists in plant cells:
![]() the plasma membrane (plasmalemma),
the plasma membrane (plasmalemma),
![]() the tonoplast,
the tonoplast,
![]() the nuclear envelope,
the nuclear envelope,
![]() Membranen des the membranes of the endoplasmatic reticulum,
Membranen des the membranes of the endoplasmatic reticulum,
![]() the membranes of the Golgi-apparatus, Dictyosomes,
the membranes of the Golgi-apparatus, Dictyosomes,
![]() the chloroplast membranes,
the chloroplast membranes,
![]() the mitochondrial membranes and
the mitochondrial membranes and
![]() the membranes of peroxysomes, glyoxysomes and other vesicles.
the membranes of peroxysomes, glyoxysomes and other vesicles.
Density gradient centrifugation is nowadays the method of choice for the separation of single compartments or membranes. In addition is the use of specific markers (for example antibodies) common that allow to find out whether a particular molecule part (an antigen) is localized at the out- or the inside of the membrane. The labelling of the antibodies with fluorescent or electron dense markers allows to detect their binding with the help of fluorescence or electron microscopes and lateral movements of the antigens within the plane of the membrane may be followed up. Apart from antibodies are lectins important as probes for the exposed sugars of glycoconjugates. They are carbohydrate-binding proteins usually of plant origin and they are able to recognize very specific sugar residues.
Most findings concerning the structural differences between different membranes were gained studying animal cells. Only during the last years were the conditions in plant cells increasingly examined. One of the reasons for this delay was that the research into animal (and especially into human) cells was done within the framework of medicinal research, especially within cancer research which is supported with huge amounts of money. Another reason is that the cell wall of plant cells proved to disrupt nearly all biochemical and molecular biological examinations. Only after it was possible to produce plant protoplasts was it possible to examine plant membranes in detail, too. Between the single compartments of a cell is constantly information, material and energy exchanged.
The plasma membrane encloses the plasma of every cell. It constitutes the interface between the cell and the extracellular surroundings. The membrane is rather thick (up to 100 Å). Besides lipids does it contain a high proportion of proteins and oligosaccharides that are usually bound to proteins but to some extend also to lipids. The majority of carbohydrates, if not all of them, is located at the outside of the plasma membrane lending the membrane an asymmetrical structure. Seen in a freeze etching specimen in top view through an electron microscope are the inner and the outer side of the membrane highly granular. In the membrane of Chlorella were particles of 80 Å diameter found that are statistically distributed over the whole membrane. Clues that they are necessary for the synthesis of the cellulose microfibrills of the cell wall exist. At the outside of the membrane is often found an unstructured layer that forms a complex with the membrane. Water and gases can pass the membrane unhindered and it is largely semi-permeable for hydrophobic substances. Apart from active transport is it essentially impermeable for hydrophilic substances.
An important property of fully differentiated parenchymatic plant cells is their extensive vacuole. It is located in the cell's centre, makes up by far the largest part of the cell's volume and is responsible for the turgor. In many other cell types, especially in meristematic tissues occur several, usually small vacuoles. Vacuoles are surrounded by a special type of membrane, the tonoplast. Both at the inside and at the outside (concave and convex surfaces) can particles of roughly 85 Å diameter be found. Vacuoles are distinguished with respect to their size, their number per cell and their contents, among which are inorganic ions, organic acids, sugars, amino acids, lipids, oligosaccharides, rubber-like substances, tannins, anthocyanes, flavonoids and more. Protein-containing vesicles of many grass and leguminous species differentiate into characteristically formed structures, the aleuron layers. Spherosomes are oil-containing particles enclosed by a lipid layer. Since a lipid layer represents - in a formal sense - just half a membrane can spherosomes strictly speaking not be counted among the vesicles.
It is generally assumed that vacuoles are temporary stores for reserve materials or final stores for waste products of the plant cell. But the stored substances of the vacuole are always of use. They help maintaining the high osmotic pressure of the cell and function as an always available nitrogen reservoir. The storage of pigments, in the petals of flowers for example, determines the flower colour and serves as a recognition signal for insects.
In some species like Hevea brasiliensis or in Papaveraceae occur numerous small vacuoles instead of just one large in latex-containing cells. Vacuoles contain usually a number of degrading enzymes (hydrolases, like proteases, exopeptidases, nucleases and other esterases). So that at least in some plant species they are attributed lysosome functions: they take part in the turnover of cellular proteins and control the turnover rates. The pH of vacuoles is generally very low, in the cells of lemon fruit, for example, is it only 2.5. This is mainly caused by the high percentage of organic acids, intermediates of the citric acid cycle that are (temporarily ?) withdrawn from metabolism. Otherwise would the pH of the cytosol become too low due to their high production rates that are a result of the hydrocarbon breakdown with the aim of yielding energy. The denaturation of many enzymes would be the consequence. Some of the organic acids of the vacuole are so concentrated that they begin to crystallize. Crystallization is particularly efficient in the presence of suitable cations like calcium (calciumoxalat formation, for example). In some Crassulaceaen species are the anions of organic acids (malate, for example) accumulated at night (CAM). They serve as carbon dioxide depots. During the day are they decarboxylated: carbon dioxide is split off and is fixed again via photosynthesis (CALVIN cycle).
A further group of substances stored in vacuoles for a fast access are amino acids. In high cytosolic concentrations would they, too, lower the pH. On the other hand are they needed as soon as the synthesis rate of certain enzymes, the growth and the multiplication of the cells increases dramatically as a consequence of external factors. Aleuron grains that can also be regarded as protein-rich vacuoles contain species-specific storage compounds, like vicelin in the cotyledons of Vicia faber.
The percentage of storage proteins in soybeans can be as high as 70% of the cell's total protein content. In vacuoles, for example in the endosperm cells of Ricinus communis, occur proteins as crystals.
One last question: How do proteins enter the vacuole, how do they pass the membrane? The answer can be found in the section about the endoplasmatic reticulum.
The nucleus is enclosed by the nuclear envelope. It consists of two membranes. It is distinguished between the outer and the inner membrane. At the nuclear pores can it be observed that the two form a continuum. Moreover exists a fluid transition between the outer membrane of the nuclear envelope and the endoplasmatic reticulum (ER). Just like the ER is it in places studded with ribosomes. The nuclear pores are filled with a pore complex of fibrillar material that takes part in the selective exchange of materials between nucleus and cytosol. At least in some species is the number of pores per square unit dependent on the activity of the nucleus. It reflects the exchange rate between nucleus and cytosol.
The genetic information is stored in the nucleus. It contains by far the largest part of the cell's DNA (the genome). Replication, transcription, RNA processing and ribosome assembly do all happen within the nucleus. The required enzymes are imported from the cytosol. Import is strictly selective and most of the cytosolic proteins are excluded. The pore complex is involved in the choice of proteins. Nuclear exports are mainly RNA and ribosomes.

The endoplasmatic reticulum (ER) is an extensive intracellular membrane system that is in direct contact with the nuclear envelope. The cytosolic surface of the membrane does often carry ribosomes (rough ER). Membrane-bound ribosomes are able to synthesize polypeptide chains for export or storage. The binding of ribosome to the membrane induces the temporary opening of a membrane pore through which the new and still unfolded polypeptide chain is synthesized. The tertiary structure forms only after entering the ER cistern. A precondition for the smuggling into the cistern is the presence of a signal sequence at the N-terminus. Cases are known, where the signal sequence is degraded after import into the cistern by specific peptidases, though exported proteins exist, where it is kept and forms an integral part of the protein structure.
Proteins are synthesized 'through' the membrane. The model shows how proteins are passed through the membrane while synthesized. The membrane pores open only after binding of a ribosome. The N-terminus of the growing polypeptide chain carries a signal- or transit sequence, the first part of the protein to be transported through the membrane. After the membrane passage is the signal sequence (usually) degraded by proteases. Different membrane types have different transit sequences (according to G. BLOBEL and B. DOBBERSTEIN, 1975).
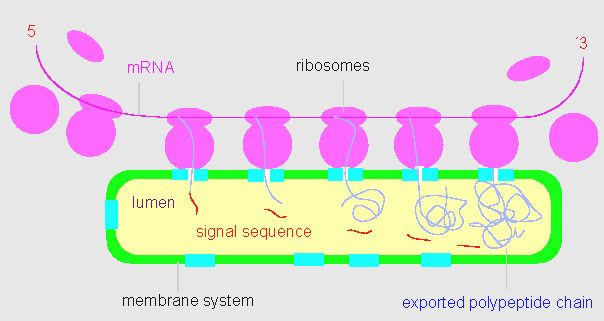
The signal sequence is specific for the membrane. Proteins that are transported through different types of membranes have an assortment of different signal sequences. Apart from the rough ER is often also smooth ER found in cells. No proteins are synthesized here, instead has the smooth ER other functions, like the synthesis of certain lipophilic substances. The endoplasmatic reticulum has to be distinguished from the Golgi-apparatus (see next section). The two organelles together form the endomembrane system.
The Golgi-apparatus is the sum of all dictyosomes and Golgi-vesicles of a cell. The dictyosomes are characteristically stacked cisterns, often branched or lobed at the margins. They carry often tubules or peripheral, small vesicles that cut off Golgi-vesicles. The dictyosomes of plant cells take part in cell wall synthesis, in the secretion of macromolecules and in the membrane flow. In the stricter sense means membrane flow the transfer of a piece of membrane from one compartment of the intracellular membrane system to another. Usually is it impossible to observe the membrane flow of a cell directly. Instead are series of electron microscopic pictures evaluated. They represent the cell's situation at a particular moment. For the reconstruction of a membrane flow have extensive picture series of cells in which the membrane flow was blocked by inhibitors to be compared with untreated control cells. Such analyses have been carried out in a big way with animal cells. They led to the conclusion that Golgi membranes can develop from the (rough) endoplasmatic reticulum. Plant data are sketchy so far but it emerges that quantitative differences in the rates of vesicular transport between the endoplasmatic reticulum and the dictyosomes exist. Moreover vary cells of different origins quantitatively.
In electron microscopic images of the cells of higher plants can it be observed that, in contrast to the situation in animal cells, the Golgi-apparatus is less closely associated with the endoplasmatic reticulum. Animal cells secrete a lot of protein, while plant cells secrete mainly hydrocarbons.
In membrane transport and in the transformation of one type of
membrane into another can at least three steps be distinguished.
- The lateral movements of lipids and proteins. Metaphorically speaking could it be said that certain molecules are selectively pushed aside.
- The fusion of two membranes and
- the subsequent budding off of vesicles.
The budding off and the fusion as well as the changes on a molecular level (membrane transformation) explain the formation of new membrane systems. Golgi vesicles can fuse with the plasma membrane and can excrete their content into the surrounding medium (called exocytosis). The reverse process occurs, too. Small parts of the plasma membrane invaginates and pinch off to form an intracellular vesicle (endocytosis). In that way are extracellular substances taken up by the cell. Two main types of endocytosis are distinguished on the basis of the size of the endocytic vesicles: pinocytosis ('cellular drinking') involves the uptake of fluid and solutes via small vesicles and phagocytosis ('cellular eating'), which involves the ingestion of large particles such as micro-organisms, cell debris or, in specialized cases, even whole cells.
To understand the export of macromolecules (proteins and poly- / oligosaccharides) are the following results decisive:
During the synthesis at the ribosomes of the endoplasmatic reticulum are the new polypeptides translocated into the ER lumen. Poly- and oligosaccharides are enzymatically synthesized from UDP-monosaccharides. The monomers are actively transported into the ER lumen or the Golgi-cisterns. The local glucosyl-transferases connect them to polymers or to oligosaccharides that are coupled to proteins or lipids (glycoconjugates).
- The pinching off of vesicles of the endomembrane system via the Golgi-apparatus is an intermediate step in the cell's secretion of macromolecules.
Though these results have mainly been gained in studying animal cells is it quite likely that the processes in plant cells are the same or are at least similar. Though the rates of synthesis and turnover may differ.
Part of the proteins synthesized through the ER membrane stay anchored in the membrane. In the case of enzymes does this mean that they may still be active inside the Golgi-vesicles and, after their fusion with the plasma membrane, at its outside. This is very important, because it offers an explanation how plant cells are able to synthesize complex polysaccharides extracellularly as in the production of cell walls and gelatinous substances.
Apart from polysaccharide synthesis and the changes and sorting of glycoproteins in dictyosomes is the intracellular transport their task. It is directed and active, that means it consumes energy. Both microtubuli and microfilaments seem to be involved.
For the fusion of Golgi-vesicles with the plasma membrane are calcium ions needed. When deleting the cell of calcium ions by chelating them with a suitable complex forming molecule occurs no fusion.
The membrane stacks of the Golgi-apparatus (dictyosomes) display a clear polarity that can be measured by a number of parameters. Quite a lot hints at a flow of cisterns through the Golgi-apparatus so that it can be spoken of a maturation of the membranes and the vesicular contents.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de