Enzymes are proteins. Every protein is determined by its amino acid sequence (its primary structure) and its tertiary structure (the three-dimensional folding of the polypeptide chain). Its uniqueness is caused by the sequence and nature of its amino acid side chains. They form a number of weak interactions, which again are the basis for the spatial arrangement (conformation) of the molecule and which help to maintain this structure by stabilizing it. In that way, the one-dimensional information that was stored in the genome as a DNA sequence is first transcribed into mRNA and after translation into an amino acid sequence finally transformed into a three-dimensional structure. It is this structure that allows an enzyme to perform its catalytic activity. That explains the specificity of catalysis and the selectivity for a certain substrate (and also that for additional regulatory factors). Enzyme molecules are, compared to most of their substrate molecules, rather large. Their surface is not evenly structured but displays dent-ins, grooves, pockets, hollows etc. The part that binds to a substrate molecule is termed the active centre or substrate-binding site and is characterized by a shape complementary to that of the substrate molecule. Furthermore, certain amino acid side chains are exposed at the active centre. They are engaged in the catalytic turn-over of the substrate.
Numerous enzymes depend on additional factors that are necessary to perform the catalytic reaction. In other words: the enzyme binds first to a coenzyme like NAD or FAD. Often, the enzyme's surface is structured in a way that the coenzyme is bound to a specifically shaped pocket. Depending on the type of molecule, the binding is either reversible (achieved through weak interactions) or irreversible (covalent bonds). The shape of the holoenzyme (= apoenzyme [protein] + coenzyme) causes the substrate selectivity. Atoms or ionized groups of the coenzyme take part in the catalytic reaction.
These properties explain, why reactions take place at an enzyme's surface that occur in solutions only after a considerable amount of activation energy has been supplied. The binding sets thr reactants into a state of enhanced reactivity recognizable by the close vicinity and the right orientation towards each other. The collision theory states that the event of two molecules meeting in solution in a way that a bond can form between them is rather improbable, though the probability can be considerably increased by the supply of energy (pressure, temperature).
As long as only a few reactants are involved, like in an experiment performed in a chemical laboratory, such methods can produce satisfying results although by-products may lower the yield. But the number of by-products would be uncountable, if the reaction started with as high a number of reactants as that of a cell - and exactly these disadvantages are avoided by enzyme catalyzed reactions. In addition, spatially and structurally separated compartments developed by the evolution of supra-molecular, complex structures, the membranes. Different sets of reactions can hence take place independent of those occurring in other compartments.
Enzyme catalyzed reactions are characterized by an intermediate enzyme-substrate complex [ES] that brings forth a product and the unchanged enzyme:
E + S > ES : k1ES > E + S : k2
ES > E + P : k3
E + P > ES : k4
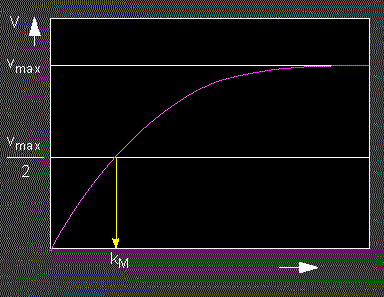
At a constant enzyme concentration, the turn-over velocity (v) of an enzyme increases as a function of the substrate concentration. It finally approaches saturation by way of an asymptotic curve. It has to be reminded that such considerations apply only to closed systems, like for example an experiment in a laboratory, but not for open systems like a plant cell where the steady state is dependent on the turn-over velocity, i.e. the efficiency of the enzyme and on a number of regulatory factors.
Velocity of an enzymatically catalyzed reaction in dependency on the substrate concentration. The yellow arrow marks the amount of substrate where the enzyme works with half of its maximal velocity: the Michaelis-Menten constant.
In the lower portion of the curve, the relation between v and
[S] , a linear one. The formation of the ES-complex as a
function of time is described by the equation
(1) d [ES] / dt = k1 ( [E] [ES] ) [S]
In a steady state, as much ES is generated as is turned over:
(2.) k1 ( [E] - [ES] ) [S] = k2 [ES] + k3 [ES]
Or, transformed:
(3) ( [E] - [ES] ) [S] / [ES] = k1 + k3 / k1 = kM
kM is also called the Michaelis-Menten constant and is an essential parameter for the characterization of a certain enzyme-substrate coupling. It is possible to calculate the concentration of the ES-complex by transformation of the equation (3) above
(4) [ES] = [E] [S] / kM + [S]
Our assumption that the starting velocity of every reaction is dependent on [ES] is expressed in the following equation:
(5) v = k3 [ES]
If we do now replace [ES] in equation (5) by its equivalent in equation (4), then we get
(6) v = k3 ([E] [S] / kM + [S])
Since the enzyme concentration [E] is rate limiting at high concentrations of substrate, is
(7 ) vmax = k3 [E]
If v is now brought into connection with vmax, then
(8) v / vmax = k3 ([E] [S] / kM + [S]) / k3 [E]
Resolved towards v
(9) v = vmax [S] / kM + [S]
This equation is known as the Michaelis-Menten equation. It describes the relation between the enzymatic rate of reaction and the substrate concentration, if vmax and kM are known. kM can be calculated, if vmax and [S] are known.
In the case that:
v = 1 / 2 vmax
is
(10) vmax / 2 = vmax [S] / kM + [S]
divided by vmax
(11) 1 / 2 = [S] / kM + [S]
or, rearranged
(12) kM + [S] = 2 [S]kM = [S]
This shows that the Michaelis-Menten constant equals the substrate concentration at half-maximal reaction velocity. Consequently, the dimension of kM is Mol. The smaller the value of kM , the higher is the enzyme's affinity for its substrate.
Since the kM can be determined from an asymptotic curve, the Michaelis-Menten equation is expressed reciprocally:
(13 ) 1 / v = 1 / vmax [S] / (kM + [S]) = kM + [S] / vmax [S]
Consequently is
(14) 1 / v = kM / vmax [S] = [S] / vmax [S]
or
(15) 1 / v = (kM / vmax) x 1 / [S]
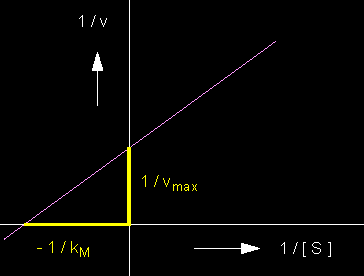
This representation is also known as the Lineweaver-Burk equation. Its advantage is in the favourable graphic depiction of the quantities. A double recessive depiction yields a straight line with a gradient of
kM / vmax
1 / kM und 1 / vmax are the points of intersection with the co-ordinates.
The dependency of the velocity of an enzymatically catalyzed reaction on the substrate concentration. Double reciprocal depiction of v and [S] according to Lineweaver and Burk
Hence, an enzyme increase the turn over velocity of a reaction, but
it has no influence on the equilibrium of the reaction itself. This
is determined by the concentrations of the reactants (law of mass
action). An enzyme has accordingly no effect on the delta G of
a reaction (the free energy), too.
|
|