|
Each cell is encircled by a membrane and most cells are equipped with an extensive intracellular membrane system. Membranes fence the cell's interior off from its surrounding. They let water, certain ions and substrates into the cell, are vital for the cell's energy budget and they excrete substances. They protect the cell. Without them would the cell contents diffuse into their surroundings, information containing molecules would be lost and the metabolic pathways would seek a thermal equilibrium: the death of each living system. |
Picture: Top left: Electron microscopic taking of an erythrocyte's membrane (plasma membrane). The membrane is seen as a double layer since electron dense material has been used for contrasting. It stains both the inner and the outer membrane's surface. The membrane is the area between the dark layers. (J. D. ROBERTSON, Duke University, Durham)
Intracellular membranes separate compartments from each other. Some surround vacuoles, others mitochondria, chloroplasts and the nucleus. Compartments are reaction areas clearly separated spatially. All biological membranes have a great deal in common as regards organization and composition:
They have an average thickness of 70 Å.
They consist mainly of lipids and proteins.
They act as a permeability barriers and are permeable only for selected substances.
When fragmented do they always reform as vesicles.
Phospholipids are the most common molecules in membranes. They consist of two parts, a hydrophilic (polar) head group (the phosphate group with the attached rest R) and a hydrophobic (lipophilic) fatty acid chain. This structure lends a tendency to form organized structures and monomolecular layers whose degree of organization is dependent on the length of the fatty acid residues and the number of double bonds to phospholipids. The Dutch E. GORTER and F. GRENDEL recognized in 1925 that two such layers of opposite orientations together would produce a fine membrane model.
The hydrophobic parts of the molecule would point towards the inside of the double layer, the hydrophilic towards its outside. In 1935 picked H. DAVSON and J. F. DANIELLI this idea up and postulated that proteins were taken up by the double layer and would interact with the hydrophilic heads of the phospholipids.
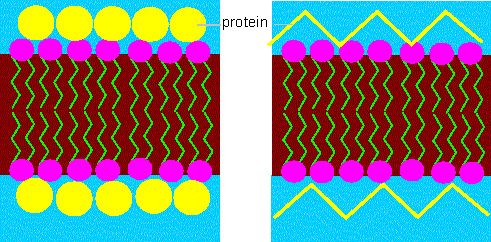
Membrane model according to DAVSON and DANIELLI. The lipids in the model form a double layer and the proteins are taken up by it. The two models (left and right scheme) are variations differing in the protein conformations.
The model went down in literature as the DAVSON - DANIELLI - model and persisted for 37 years. It explains the organization of the lipid molecules but could not account for numerous properties of membrane proteins without doubt.
In 1972 was it superseded by the fluid mosaic model of J. S. SINGER and G. NICHOLSON (University of California, San Diego).
The model describes the membrane as a fluid mosaic. It suggests
that the lipids form a viscous, two-dimensional solvent into which
proteins are inserted and integrated more or less deeply.
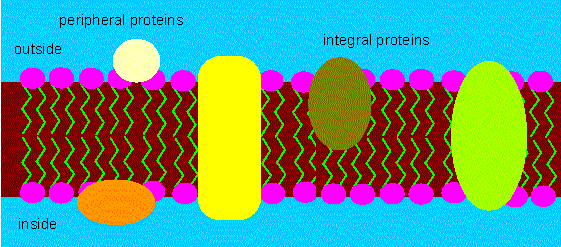
Fluid mosaic model of a membrane. The proteins are inserted into the double layer and are not merely attached to it. Some of them span the whole membrane's thickness. The hydrophobic parts are in contact with the hydrophobic tails of the lipid molecules. The polar groups jut out from the membrane. (according to S. J. SINGER and G. L. NICOLSON, 1972).
Proteins of this category are called integral membrane proteins. Some of them stretch from side of the membrane to the other, many are joined in aggregates and it can easily be imagined how channels and pores through the membrane form. The model states also that the molecules can move laterally and freely within the membrane. This seems to contradict the observation that certain molecules are often not statistically distributed and that the membrane is, like a mosaic, organized into areas of different functions. This can be accounted for by protein-protein interactions that restrict the movements of the single molecules. In addition is it known that peripheral proteins attached at the outer or inner surface exit beside the integral ones. They may also interact with each other. Protein parts in contact with the aqueous phase are often glycosylated.
| Porin from Rhodobacter capsulatus | Porin from Rhodpseudomonas blastica | PhoE from Escherichia coli |
Exchange of molecules through membranes depends on highly specific transport systems, prominent representatives of this group are the porines that extensively have been studied in bacteria. In the picture series above porines from several species are shown to illustrate that these proteins are highly conserved in evolution. The figures look rather transparent, this, however, is a wrong impression. Use right mouse key to click in figure frame for additional features of the molecular model. Choose the options display/spacefill. - A few more examples of pores and channels are shown in the interactive files below:
The specific form of some cells (erythrocytes, for example) is determined by such a protein skeleton that spans the membrane.
The single membrane types of a cell differ due to different compositions:
Different phospholipids (and other lipids, like cholesterol) in the two layers cause an asymmetry. The differences can be of a qualitative or quantitative nature.
Membranes of different origin differ also in their lipid compositions
Different proteins cause different functions of membranes. Some enzyme activities can be used as markers for certain types of membranes.
The orientation of some proteins is important, too.
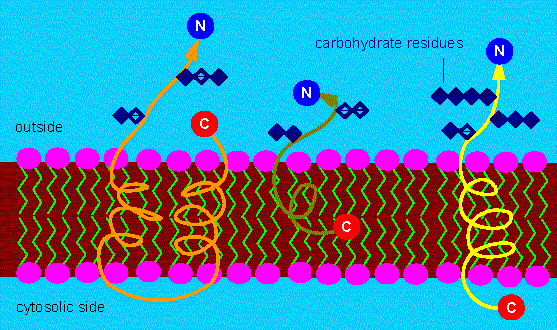
Different possibilities to fix integrated membrane proteins within the membrane. The N-terminus is always at the side that is turned away from the cytosol. Often are hydro-carbon residues attached to the N-terminus of proteins. Typical membrane proteins can therefore be counted among the glycoproteins (exceptions exist). The C-terminus can either be at the outside, within the lipid layer or protrude into the cytosol (the site of protein synthesis).
Membranes have numerous functions. The criteria mentioned above can be further distinguished:
Permeability for one type of molecule and impermeability for another belong to the properties that hint at the presence of a membrane. All biological membranes are selectively permeable.
Membranes separate reaction areas that can be distinguished due to their chemical composition, their pH value and their electric potential. Both the pH and the electric gradient can be measured.
Some ions or small molecules (substrates) can be transported through the membrane. This transport is energy-consuming. It is also spoken of an active transport in contrast to passive transport that occurs only in the direction of the concentration gradient.
Due to different compositions have the single membrane types different functions, too. Highly specialized membranes like the thylacoid membranes of chloroplasts and the inner membranes of mitochondria are involved in energy turnovers.
In order to understand passive transport is it necessary to understand a little of the topics diffusion, permeability and osmosis, too. Active transport can be understood when knowing structure and function of integral proteins and the electrochemical properties of the membrane while the understanding of energy transformation requires information about all the molecules within the respective membrane.
Related questions are: which reactions take place within the different compartments, which importance has the spatial separation of single biosynthetic pathways, how does the substance - and energy exchange between the single compartments take place? How is information exchanged? How are the different ion - and metabolite concentrations maintained? How are large molecules (like proteins) transported through the membrane and, finally, how are membranes made? Can they change into each other and how would such a membrane flow look like? Which influence does it exert on the molecular transport within the cells, from cell to cell and between the cells and their environment?