
A number of plants display an increased and more efficient net photosynthesis during strong light intensities. A prime example are the Gramineae of warmer regions like maize or sugar-cane.
At the beginning of the sixties observed H. KORTSCHAK (Hawaiian Sugar Planter's Association) that the first product of photosynthesis in sugar-cane is not the C3 unit 3-phosphoglycerate but a unit with four C-atoms. The Australian plant physiologist M. D. HATCH and his English colleague C. R. SLACK confirmed this result and identified the compound as oxaloacetate (OAA). It is produced by the addition of one molecule of carbon dioxide to phosphoenolpyruvate (PEP). The cycle is also known as the HATCH- SLACK-cycle or the C4 cycle. Plants with this cycle are called C4-plants (and CAM plants, respectively) in contrast to C3 plants where the carbon dioxide is directly fed into the CALVIN cycle. The oxaloacetate is usually converted into malate of which the carbon dioxide is split off again with the help of an enzyme.
This carbon dioxide is now bound by ribulose-1,5-diphosphate and assimilated via the CALVIN cycle. :
Some species use malate instead of aspartate
oxaloacetate + L-glutamate > aspartate + alpha-ketoglutarate.
The reversible binding of carbon dioxide has the function to accumulate and store CO2. The process consumes energy, so that it could also be spoken of a carbon dioxide pump. It should be mentioned that the HATCH-SLACK cycle requires two molecules of ATP are per fixed carbon dioxide.
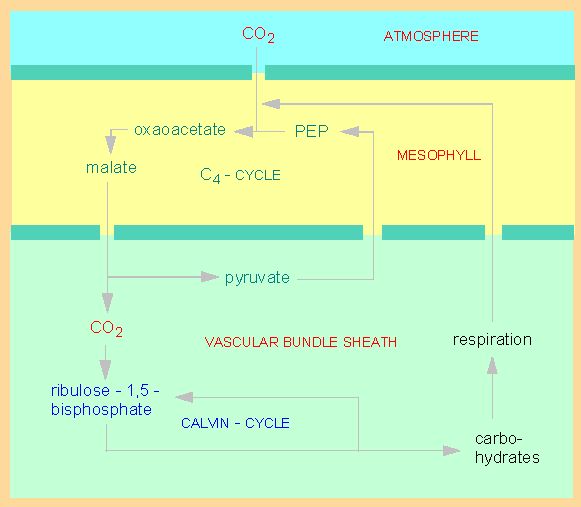
Photosynthesis of C4 plants. CO2 is bound to phosphoenolpyruvate (PEP) in mesophyll cells. The product is oxaloacetate. The next step generates malate. In the cells of the vascular bundle sheath, the 'Kranz' cells, is carbon dioxide split off the malate and fed into the CALVIN cycle. The pyruvate is transported back into the mesophyll cells (active transport) and is with the help of additional ATP phosphorylated to PEP.
The anatomy of C4 leaves with so-called 'Kranz' cells differs fundamentally from that of C3 plants. The chloroplasts of C3 plants are of homogeneous structure, while two types of chloroplasts occur in C4 plants. The mesophyll cells contain normal chloroplasts, that of the vascular bundle sheath have chloroplasts without grana , i.e. they are partially impaired in function. This peculiarity does not affect the CALVIN cycle, it concerns only the light reactions of photosynthesis. The first binding of carbon dioxide (the HATCH-SLACK reaction) occurs in the mesophyll cells, the incorporation into carbohydrates (the CALVIN cycle) in the cells of the vascular bundle sheath. Both processes of photosynthesis are spatially separated.
CAM is the abbreviation of Crassulacean acid metabolism. The name points at the fact that this pathway occurs mainly in Crassulacean species (and other succulent plants). The chemical reaction of the carbon dioxide accumulation is similar to that of C4 plants but here are carbon dioxide fixation and its assimilation not separated spatially but in time. CAM plants occur mainly in arid regions. The opening of the stomata to take up carbon dioxide is always connected with large losses of water. To inhibit this loss during intense sun (the transpiration via the cuticle remains intact) has a mechanism developed that allows the uptake of carbon dioxide during the night. The prefixed carbon dioxide is stored in the vacuoles as malate (and isocitrate) and is used during the daytime for photosynthesis.
The enzyme that catalyzes the primary carbon dioxide fixation
of C4 and CAM plants is phosphoenolpyruvate carboxylase (PEPC).
Its affinity for carbon dioxide is by far higher than that of
Rubisco, the first enzyme of the CALVIN cycle. As a consequence
are C4 plants able to use even trace amounts of carbon dioxide.
PEPC occurs in small amounts (roughly 2 - 3 %) also in C3 plants,
where it, too, has a key position in the metabolic regulation.
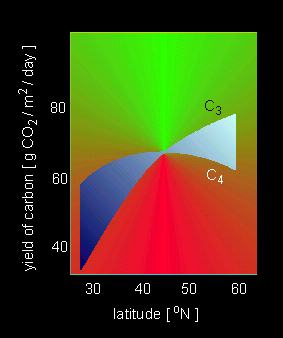
Carbon dioxide yield of C4 and C3 plants of open grasslands in different parts of the world. In temperate regions is the rather low light intensity decisive for the disadvantage of C4 plants. C3 plants have an advantage due to their low rate of photorespiration and because they need no energy for the previous fixation of CO2. (J. R. EHRLICHER, 1978).
In growing roots (of maize seedlings, for example) does PEPC help to supply the lipid synthesis with NADH + H+. The following reactions take place:
- phosphoenolpyruvate + HCO3- > oxaloacetate + Pi
- oxaloacetate > malate. - During this step is NADH + H+ oxidized to NAD+.
- malate > pyruvate + CO2.
During the last reaction is NAD reduced to NADH + H+.
In the root nodules of leguminosae is nitrogen fixed. Enough carbon bodies have to be supplied in order to incorporate the ammonia produced by the bacteria. the CO2-binding via PEPC-reaction thus is an important supplement
Furthermore produces the PEPC intermediates of the citric acid cycle (oxaloacetate and / or malate), a back-up in case of shortages. The activity of PEPC is controlled by extern factors, the day length is decisive. In some cases have different isoenzymes be found in different tissues. Their production is controlled by different triggers.
C3 plants can loose up to 20 percent of the carbon fixed in the CALVIN cycle at intense radiation. Under strong light is the photorespiration 1.5 - 3.5 times as high as the usual respiration in darkness. In C4 plants becomes the photorespiration drastically reduced, it may even not be detectable any more. In other words:
The net rate of photosynthesis (and consequently also the net production of biomass) of C4 plants is far larger at high light intensities than that of C3 plants. The optimal temperature of photosynthesis is below that of the respiration in darkness. As a consequence are losses caused by respiration larger at high than at low temperatures. Where light is a limiting factor and temperatures are low (i.e. in temperate climatic zones) have C3 plants the advantage, C4 plant do hardly occur (one of the exceptions is Spartina townsendii). C4 plants, nearly always herbs or shrubs, are more successful in the open country of warmer zones.
It has to be mentioned that two molecules of ATP are consumed in the HATCH-SLACK cycle
C4 plants belong to numerous, phylogenetically not related monocotyledonous and dicotyledonous families. Moreover have C4 activities also been detected in the blue-green alga Anacystis nidulans as well as in some dinoflagellates.
Since the alternative C3 or C4 is accompanied by considerable changes of the leaf anatomy has it to be assumed that the genetic potential for both pathways is quite common in the plant kingdom and that, depending on the ecological needs, one way is chosen by a species while a related species may choose the other one.
A well-studied example is the genus Atriplex, where both ways are realized. The C3 plants belong to one phylogenetic group, the C4 plants to another. In some cases can hybrids of C3 and C4 species be generated.
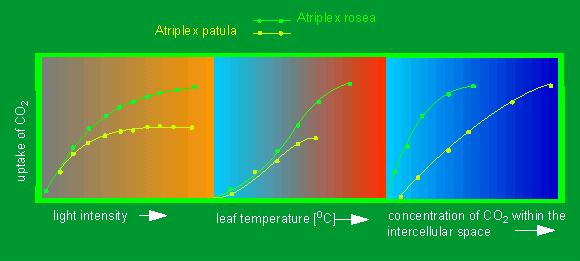
Influence of different parameters on the efficiency of the carbon dioxide uptake (ordinate) of a C3 plant (Atriplex patula, yellow line) and a C4 plant (Atriplex rosea, green line). Measured parameters (from left to right): light intensity, leaf temperature and concentration of carbon dioxide within the intercellular space (according to O. BJÖRKMAN and J. BERRY, 1973).
In several plant species of the genera Zea, Mollugo, Moricandia, Flaveria, etc. occur both types of CO2 fixation within one plant. In younger plants is usually the C3-, in older ones the C4 pathway taken. The amount of C4 is controlled by environmental factors.
CAM: Advantages and Disadvantages. CAM has been detected in more than 1000 angiosperms of 17 different families. It is usually accompanied by succulence, though not all Crassulaceae, for example, display CAM and succulence is no precondition of CAM. Tillandsia usneoides of the bromelia family is not succulent, but uses CAM. Mesemryanthemum crystallinum (a plant with succulent leaves) can use the C3 pathway but switches to CAM when growing in saline soils. Under experimental conditions can the shift be achieved by increasing the NaCl concentration of the nutrient medium (K. WINTER and D. J. von WILLERT, 1972). While the advantage of C4 plants comes in useful under high light intensities, is the degree of the CAM influence in CAM plants regulated mainly by temperature, atmospheric humidity and salinity. Both strong and weak CAM plants are known. In weak CAM plants becomes CAM only apparent at certain differences between day and night temperature. CAM plants that store a lot of malate and due to the thus high osmotic value also a lot of water, are usually less frost resistant than C3 plants. Because of the high concentration of acid are they less heat resistant, too. Species of arid regions are therefore forced to break their pool of malate down during the daytime (R. LÖSCH and H. KAPPEN, Universität Kiel, 1985). Usually do the C4 pathway and CAM exclude each other. An exception is the succulent C4 dicotyledon Portulaca oleracea that is able to choose the optimal pathway. under natural conditions
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de