

Viruses are pathogens with an extremely narrow host range. Their phylogenetic origin is vague, tough it has always been considered that viruses are genes that became vagrant after having excluded themselves of the host’s or a related species’ genome. During the last years, an alternative explanation assuming viruses to be by-products of RNA processing has been discussed. All suggestions remain hypotheses, though, since no definite proof for either of them exists yet. Detailed descriptions of all plant viruses can be found in the data base VIDE (Plant Viruses Online), that is accessible, for example, via the Botany online interface:
 |
Viruses (Virus particles or virions) are usually units consisting of nucleic acids and coat proteins called capsids. Viroids consist only of RNA, i.e. they contain no protein at all. Except for a few cases, viruses are not surrounded by a membrane. If present, the membrane around a virusparticle - as seen in electron microscopic images - stems usually from the host cell (see picture to the left). Viruses have no energy metabolism of their own. Consequently, they cannot perform syntheses and are thus unable to replicate themselves. Depending on their host species, it is distinguished between plant viruses multiplying almost exclusively within plant cells, bacterial viruses (bacteriophages) that depend on living bacteria, and animal viruses.
|
With plant viruses, the term specificity (or host-specificity) has a very narrow meaning, since no plant virus as such exists. Instead, plant viruses can be grouped in a number of ‘varieties’. The tobacco mosaic-virus (TMV), for example, multiplies within Nicotiana-species, several other solanaceous plants, and a few species of other plant families. The name of a virus is usually derived from the name of its main host plant. Although with viruses, the term ‘species’ may not quite correspond to the way it is defined in biological systematics, it is perfectly reasonable and common to use it for viruses, too, since all viruses and viroids contain an original genome with a species-specific information. Its continuity over generations is guaranteed by replication in the host cells. The genetic information of viruses is either encoded by single-stranded RNA (most plant viruses), double-stranded RNA (wound tumor viruses), single-stranded DNA (gemini-viruses) or double-stranded DNA (cauliflower mosaic-virus: CaMV). Based on the shape of the virus particle, it is distinguished between rod-shaped and icosaedrical viruses with a capsid that seems almost spherical.

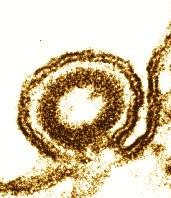
Tobacco mosaic virus:
Electron micrograph of a negatively stained specimen. The relief-like structure is a consequence of image processing.
Plant viruses have no specific mechanism for entering the host cell. Cell wall and cuticle are difficult obstacles for them. Plant viruses depend therefore on injuries or on transmission via invertebrates (insects, nematodes, etc.). The animal transmitter does in some cases also act as an intermediate host. This means that some plant viruses are able to multiply within animal tissue.
Virus diseases of plants are relatively rare. Infection is scarcely strong enough to kill the plant. Monoculture favours spreading, and agricultural losses of profit caused by the potato-X, or the potato-Y viruses, for example, can be significant.
Of many viruses, numerous, considerably differing strains (wildtypes) have been isolated. Among the differences are host range and the degree of virulence. Moreover, viroids were detected. Viroids are small, circular RNA molecules that do not encode proteins themselves. Instead, virions interfere with the transcription of cells due to their similarity with certain areas of recognition of primary transcription products. It seems that viroids prevent the correct cutting out of the introns. They are presumably multiplied with the aid of the cellular DNA-dependent RNA-polymerase II. Viroids occur mainly in warm climates and cause significant loss of profit as the causative agents of the potato disease or the Cadang-Cadang disease of palms.
The virus concentration within plant cells is high, although a virus like the TMV does not harm the host seriously. Infected cells contain often voluminous virus crystals. TMV was thus the first virus to be isolated in a pure state and in large amounts (W. M. STANLEY, 1935, then at Princeton University).
Plants are far from being defenceless against viruses. Only a few virus species are able to penetrate meristematic tissues or to infect a number of successive plant generations (vertical transmission). Hypersensitivity is an effective protection. We know it already from fungus infections. It is based on the dying of cells in the immediate surrounding of the primary site of infection, thus stopping the spreading of the virus. In the case of Nicotiana tabacum, the genetic cause of hypersensitivity has been analyzed. One Nicotiana gene product guarantees hypersensitivity against all TMV strains, while another affects only some TMV strains. The symptom a virus causes at the primary site of infection is called primary symptom. Symptoms caused by its spreading throughout the rest of the plant are called secondary symptoms .
Virus infections can usually be recognized by mosaic-like leaf patterns of light and dark green. The infection spreads often over the whole leaf beginning at the leaf veins. Leaves that had been infected during their development are usually deformed or involute.
Frequently, lightened leaf areas, called chloroses, develop around the primary site of infection. Withered areas are called necroses . Chloroses are caused by a breakdown of the chlorophyll resulting in a decreased rate of photosynthesis. Heavy infections are characterized by a complete local loss of chlorophyll. Affected areas have a yellowish look as only the carotenoids remain. Some TMV strains, for example, can be recognized by yellow leaf areas ("yellow strains"). In nature, such strains are pretty rare since such a strong damaging of the host does also impair the virus’ chances of replication and spreading.
Some viruses multiply within the plant without causing symptoms. This phenomenon is called latent infection. In contrast, wound tumor virus cause the development of tumors. The symptoms of most viruses are dependent on both virus and host, and do thus present an important diagnostic feature.
G. SAMUEL proved in 1934 experimentally that the TMV virus uses the plant’s vascular system for spreading, actually both the plant’s transport to and away. As a result, fully differentiated, old leaves and roots, and young leaves are equally infected.
Since decades now, plant viruses (and especially the TMV) are favourite test objects of pure research due to the large amounts of material extractable from infected plants. The TMV was, too, the first biological electron-microscopic object to be photographed (G. KAUSCHE, E. PFANNKUCH, A. RUSKA, Berlin, 1939).
Isolated TMV-RNA itself is infectious. It can thus be a carrier of genetic information (A. GIERER and G. SCHRAMM, Tübingen, 1956). Chemical changes of the TMV-RNA (deamination of adenine or cytosine due to treatment with nitrite) showed in addition and for the first time that changes of the RNA are mutagenic, too (K. W. MUNDRY and A. GIERER, 1958). The changes of these first experiments caused mutations of a coat protein (A. TSUGITA and H. FRAENKEL-CONRAT, Berkeley, 1960; H. G. WITTMANN, Tübingen, 1960).
The analysis of a large number of chemically induced mutants (performed by the working groups above (TSUGITA and FRAENKEL-CONRAT, WITTMANN)) yielded the first results important for the encoding of the genetic code. WITTMANN’s data proved that the genetic code is not overlapping, and that it is universal in all biological kingdoms: bacteria, plants, and animals. Work on the TMV made plain, too, that the gene-carrying RNA does not simultaneously function as mRNA. mRNA is synthesized within the cell as a separate fraction (T. R. HUNTER et al., Cambridge, 1976).
The working group of A. KLUG (Medical Research Council, Laboratory of Molecular Biology, Cambridge) studies the assembly of complete virus particles from protein subunits and the incorporation of RNA into the newly formed virus particle since more than two decades.
The development of the X-ray analysis (Röntgen structure analysis) and the quicker examination of data due to a new generation of computers allowed the clearing up of the viruses’ tertiary protein structure., The substantial advance of the studies reviewed here piloted S. HARRISON and his colleagues (Harvard University) in the early eighties to the analysis of spherical viruses. Their studies resulted in the unravelling of the viral tertiary and quaternary structure. The minimum limit of resolution of 2.8 to 2.9 Å was sufficient for the localization of a single amino acid.
Semi-equivalent aggregation was an important discovery in these studies. The coat protein of some viruses is able to aggregate into different forms. The forms differ in their number of symmetry axes and in their molecular assembly. The generation of such aggregates explains an as complex structures as a hollow sphere. The findings of HARRISON et al. have model character, since complex structures of this type are frequent in nature. The screw-like flagellum of prokaryotes, for example, is an aggregation of a single type of protein (flagellin).
So why have molecular biologists studied bacteriophages instead of plant viruses so intensely and for decades? Here are some plausible answers:
The multiplication of bacteriophages is easier to understand. Their hosts, the bacteria (Eschericia coli has been studied best) are easy to cultivate, the duration of one generation is short. A complete circle of the phages’ multiplication is finished within 15 minutes. The phage / bacterium system is less complex than the virus / plant system. The single parameters are thus easier to measure. Infection with a single phage is sufficient, while in the case of plant viruses often several viruses have to participate.
Geneticists studying bacteria and bacteriophages were mainly interested in the (double-stranded) DNA of the phages (RNA-containing phages and single stranded DNA phages exist, too). The information of DNA double-strands can be recombined, the order of its genes is determinable. RNA molecules have no recombination, it is therefore often difficult to say something about the organization of the plant viruses’ genomes. Modern genetic engineering, nevertheless, offers possibilities to obtain results. A disadvantage of the phages is the frequently complex structure of their capsids. The coat consists often of more than 20 different proteins.
The study of bacteria and their bacteriophages yielded numerous results regarding the course of the infection and the biosynthesis of bacteriophages. Experiments showed the astonishing performances of cells. During the late fifties, for example, mRNA was detected within a phage-infected bacterium. Later on, mRNA was found to be an essential intermediate of every protein biosynthesis.
The study of phytopathogenic viruses raised hopes of obtaining knowledge about the possibilities and the expression of the plant genome, as well as the defence mechanisms of a plant against foreign RNA or DNA.
Based on the results of molecular biology, and especially on the virus research of the last decades, it was tried to develop a protection against virus infections with the help of the now available methods of genetic engineering. An example is the attempt to induce a defence mechanism against the leaf-curl virus. A report from W. SCHUCHERT, E. PEERENBOOM, and the MPI für Züchtungsforschung (in German only) can be found under:
|
|