Die Abteilung Phaeophyta enthält nur eine Klasse, die Phaeophyceae (=Fucophyceae), die in 11-13 Ordnungen untergliedert wird. Es gibt etwa 1500 bis 2000 Arten, die 250 Gattungen zugeordnet werden. Sie kommen fast nur im marinen Bereich vor. Wie die Rotalgen bevorzugen sie die Litoral- und die Sublitoralzone. In klaren tropischen Gewässern wurden sie noch in Tiefen von 200 Metern nachgewiesen. Ihr Hauptverbreitungsgebiet sind die Meeresküsten gemäßigter Zonen. Braunalgen können eine beträchtliche Größe erreichen (Macrocystis und Nereocystis bis etwa 100 m Länge); die großen Formen werden als Tange bezeichnet. Durch spezielle Haftorgane sind sie in der Regel an feste Unterlagen (Fels, Muschelschalen usw.) fixiert (benthische Lebensweise). Wenige Arten (z.B. Sargassum) kommen im freien Oberflächenwasser wärmerer Meere (Sargassosee) treibend (pelagisch) vor. Die Gameten der Braunalgen sind meist vielgeißlig. Eizellen und Tetrasporen hingegen sind - sofern vorhanden - unbegeißelt
Auf zellulärer Ebene gibt es eine Menge von Gemeinsamkeiten mit den Chrysophyceae, den Bacillariophyceae und den Xanthophyceae:
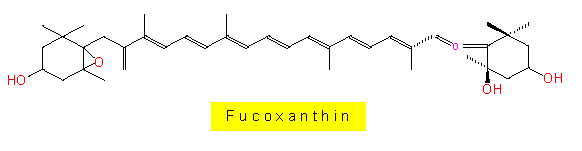
| ähnliche oder fast gleiche Pigmentzusammensetzung (Chlorophyll a und c, Fucoxanthin [außer bei Xanthophyceen]), | |
| nahezu identische Feinstruktur der Plastiden, | |
ungleich lange (heterokonte) Geißeln, die aber immer seitlich ansitzen und von denen eine mit zwei Reihen dreiteiliger steifer Haare (Mastigonemen) besetzt ist. Die andere hat, zumindest bei den Zoosporen, eine Geißelanschwellung, die als Photorezeptor dient. |
|
| Augenflecken sind, wenn vorhanden, in Plastiden eingeschlossen. | |
| Ähnlichkeiten im Bau und in der Bildung des Golgi-Apparats, | |
| Reservepolysaccharide werden an Pyrenoiden gebildet. |
Aufgrund dieser Merkmale sind die genannten Gruppen verschiedentlich einer gemeinsamen Abteilung (Heterokontophyta) zugewiesen worden. Die aufgeführten Gemeinsamkeiten sind stichhaltige Beweise für einen monophyletischen Ursprung. Es ist daher auch unbestritten, daß sich alle (auch die Braunalgen) von einzelligen Arten ableiten, die sich durch die obengenannten Merkmale ausgezeichnet haben müßten. Die gemeinsame Herkunft rechtfertigt jedoch nicht unbedingt den Zusammenschluß von Taxa zu einer Abteilung. Gerade während der Evolution des vielzelligen Braunalgenthallus traten Eigenarten in Erscheinung, die auf der Organisationsstufe Zelle noch nicht vorkamen. Keine der Besonderheiten der Architektur des Braunalgenthallus kommt bei einem der anderen hier genannten Taxa vor. Vergleichbare Baupläne findet man allenfalls bei den ihnen phylogenetisch weniger nahestehenden Rot- und Grünalgen.
An anderer Stelle werden wir die weitgehenden Übereinstimmungen im Bau und in der Art der Fortpflanzung der Pteridophyten und der Spermatophyten herausarbeiten. Niemand zweifelt daran, daß sich letztere von fossilen Pteridophyten (Vorfahren der Samenfarne) ableiten. Trotzdem besteht eine einhellige Meinung darüber, die beiden Gruppen als eigenständige Abteilungen zu behandeln. Praktische und traditionelle Gründe spielen hierbei ebenfalls eine wichtige Rolle.
Die Ordnungen der Phaeophyta können aufgrund zunehmender Komplexität im Thallusbau, der ontogenetischen Differenzierungsweise, der Akzentuierungen in ihrem Lebenszyklus und der Spezialisierung im Bereich der vegetativen und generativen Fortpflanzung charakterisiert werden.
Als ursprüngliche Ordnung gelten die Ectocarpales. Unter Beachtung bestimmter Kriterien läßt sich zeigen, daß im Verlauf der Braunalgenevolution drei voneinander unabhängige Wege beschritten wurden:
Beibehaltung des fädigen Thallus, aber Entwicklung morphologischer Unterschiede zwischen Sporophyt, männlichem und weiblichem Gametophyten: Chordariales, Desmerestiales, Sporochnales. |
|
Entwicklung komplexer Thalli mit erkennbarer Gewebedifferenzierung, männliche und weibliche Gametophyten unterschiedlich gebaut: Scytosiphonales, Dictyosiphonales, Cutleriales, Laminariales. |
|
Entwicklung komplexer Thalli mit erkennbarer Gewebedifferenzierung. Beibehaltung gleichaussehender männlicher und weiblicher Gametophyten: Fucales, Dictyotales, Sphacelariales. |
Einige der Besonderheiten der Entwicklungstendenzen sollen an den vier folgenden Beispielen illustriert werden (für Beibehaltung des fädigen Thallus, Unterschiede zwischen männlichem und weiblichem Gametophyten wird kein Beispiel gebracht).
Bau und Fortpflanzung von Ectocarpus siliculosus. Einzelheiten wurden von D. MÜLLER (Universität Konstanz) an Material aus Neapel erarbeitet. Der wenige Zentimeter große Thallus besteht aus verzweigten Fäden. Diese verlängern sich durch interkalare Teilungen (also kein Spitzenwachstum !). Die Sporophyten (2n) ähneln den Gametophyten (1n), die allerdings meist etwas stärker verzweigt sind. An den Enden kurzer Seitentriebe werden Sporangien angelegt. Dabei lassen sich vielkammrige von einkammrigen unterscheiden. In den vielkammrigen entstehen diploide Zoosporen, aus denen sich wieder ein Sporophyt entwickeln kann (ungeschlechtliche Fortpflanzung). In einkammrigen erfolgt eine Meiose; im Anschluß daran eine Reihe weiterer mitotischer Teilungen, schließlich werden zahlreiche haploide Gameten freigesetzt. Aus je etwa der Hälfte entwickelt sich ein weiblicher bzw. ein männlicher Gametophyt. Es sieht demnach ganz so aus, als sei die Geschlechtsbestimmung genetisch determiniert. In den vielkammrigen Gametangien der (zweihäusigen) Gametophyten entstehen die beweglichen männlichen bzw. weiblichen Gameten. Die männlichen Gameten finden die weiblichen aufgrund chemotaktischer Anlockung, denn jene sondern ein Gamon (einen Sexuallockstoff) ab. Seine chemische Struktur sowie die der Gamone weiterer Ectocarpus-Arten und anderer Braunalgen ist bekannt. Sie alle gehören einer Molekülklasse an, unterscheiden sich aber deutlich von Art zu Art (L. JAENICKE, Institut für Biochemie, Universität zu Köln, 1977). Aus der Zygote entwickelt sich wieder ein diploider Sporophyt. Die Chromosomenzahl von Ectocarpus siliculosus liegt bei 2n = 50.
Sphacelaria plumosa. Typisch sind eine Scheitelzelle, sowie die Entstehung primärer und sekundärer Segmente. Eine Gewebebidung erfolgt durch Längsteilungen. Spezialisierte Zellen (Phäophytenhaare) treten auf. Vegetativ vermehren sich die Sphacelariales durch Brutkörperbildung. Bei der generativen Fortpflanzung sind Anisogamie und ein isomorpher Generationswechsel nachgewiesen worden.
Dictyota dichotoma (Gabeltang). Über diese Art und die Entstehung ihres dichotom verzweigten Thallus haben wir bereits gesprochen. Er ist ein klassisches Objekt botanischer Anfängerpraktika. Hier erfolgt das Wachstum durch Teilungen einer apikal sitzenden Scheitelzelle, deren Fähigkeit sich längs zu teilen Ursache der echten dichotomen Gabelung ist. Dictyota dichotoma ist an südeuropäischen Küsten - sofern Felsen vorhanden ist - häufig. Früher fand man sie auch regelmäßig im Helgoländer Felswatt, heute nur noch sporadisch. Ein Grund dafür ist vermutlich eine Temperaturabsenkung um wenige Grade, die in den letzten Jahrzehnten registriert wurde. Für Dictyota lag Helgoland schon früher an der Nordgrenze ihrer Verbreitung.
Da Sporophyt und Gametophyten gleichgestaltet sind, die Kernphasen sich aber unterscheiden, haben wir hier einen typischen isomorphen, heterophasischen Generationswechsel vor uns. An einzelnen Rindenzellen der Sporophyten werden in lockerer Anordnung kugelige Tetrasporangien gebildet, in denen durch Meiose jeweils vier haploide geißellose Tetrasporen entstehen. Aus ihnen entwickeln sich häufig männliche und weibliche Gametophyten (genotypische Geschlechtsbestimmung).
Auf dem männlichen Gametophyten entstehen sogenannte "Antheridien", die den plurilokulären Gametangien der Ectocarpales homolog sein dürften. Sie sind zu deutlich umgrenzten Gruppen auf der Thallusoberfläche, den sogenannten "Sori" zusammengefaßt. Die aus ihnen freigesetzten Gameten besitzen nur die vordere pleuronematische Geißel. Auch die Gametangien der weiblichen Gametophyten sind zu Sori zusammengefaßt, die (anders als die männlichen Sori) an ihrem Rande von sterilen Zellen umgeben sind. Die einzelnen Gametangien enthalten jeweils nur einen einzigen Gameten, der geißellos ist. Wir haben es hier also mit Oogamie zu tun
Die Eizellenfreisetzung folgt einer lunaren Rhythmik. Die Sori
werden während der Nipptide angelegt, und die Gameten werden
an Tagen nach der folgenden Springtide frei.
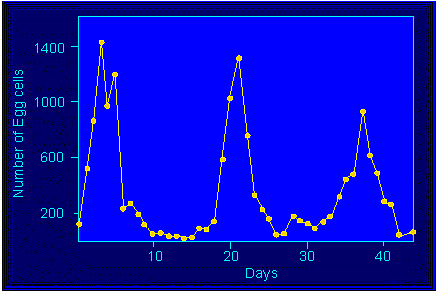
Dictyota dichotoma: Entleerung von Eiern aus den Oogonien unter Laborbedingungen (Lichtprogramm: 14 Stunden Licht, 10 Stunden Dunkelheit). Ordinate: Anzahl der an den betreffenden Tagen gezählten freigesetzten Eizellen (E. BÜNNING und D. MÜLLER, 1962).
 Laminariales
Laminariales
Zu den Laminariales gehören die größten
Tange. Im Bereich des Kontinentalschelfs bilden sie ausgedehnte
Wälder (oder Wiesen). Solche Wiesen kommen u.a. im Felswatt
vor Helgoland vor. Vor der nordamerikanischen Pazifikküste
bestehen die Wälder aus riesigen Exemplaren der Gattung Nereocystis;
in der südlichen Hemisphäre wird sie durch Macrocystis
vertreten. In der Antarktis ist die Gattung Lessonia mit
einem Thallus von palmähnlichem Habitus vorherrschend.
Der "typische" Laminarienthallus (Sporophyt !) ist dreiteilig und besteht aus:
| einem meist krallenförmig gebauten, vielfach verzweigten Haftorgan(Haftkrallen, Rhizoide), | |
| einem oft viele Meter langen seilartigen Stiel (Cauloid), und | |
| einem flächigen, oft unterteilten Thalluslappen (Phyloid). |
Bei vielen Arten sind voluminöse, innen hohle Schwimmkugeln ausgebildet, die z.B. bei Nereocystis Durchmesser von 15 bis 20 Zentimetern erreichen und Kohlenmonoxyd (!) enthalten. Die auch an europäischen Küsten verbreiteten Laminaria-Arten sind meist mehrjährig, wenige (z.B. Laminaria ephemera) sind einjährig. Während der Überwinterung perennierender Arten bleiben vielfach nur die Haftkralle und das Cauloid erhalten, das Phyloid wird im Frühjahr regeneriert. An ihren oberen Enden sind Thalluslappen unregelmäßig zerfranst. Teilungsfähiges Gewebe ist stets an der Basis in einem interkalaren Meristem lokalisiert. Dadurch wird neues Gewebe nachgeschoben; die an den Enden sitzenden, durch Wellengang abgenutzten Zellen werden somit periodisch ersetzt. Die Form der Thalluslappen ist ein artspezifisches Merkmal. Experimentell - d.h. durch Verpflanzungsversuche - ließ sich jedoch zeigen, daß sie durch Umweltfaktoren (Strömung, Wassertiefe, Licht, Temperatur) in weiten Grenzen modifizierbar ist. Laminaria digitata z.B. bildet in strömendem Wasser schmale Thalluslappen aus; nach Übertragung an geschützte Standorte entstehen breite, dicke Thalli, wobei die Dickenzunahme auf der Ausbildung zusätzlicher Zellschichten beruht. Die morphologisch erkennbare Differenzierung spiegelt die auf Zellebene erkennbare Differenzierung in Gewebe wider. Wie bei den Fucales (s. folgenden Abschnitt) können corticale und medullare Gewebe voneinander unterschieden werden. Die Photosynthese erfolgt zum überwiegenden Teil im corticalen Bereich. Die wichtigsten Reservestoffe sind das Laminaran (ein beta- 1,3 -Glucan), der Alkohol Mannitol und Öle. Der Transport dieser und anderer Substanzen (z.B. Aminosäuren) erfolgt in spezialisierten leitbündelähnlichen Geweben, die aus Bündeln langgestreckter, an den Querwänden siebartig durchbrochener Zellen bestehen. Damit ähneln sie sowohl in der Form als auch in der Funktion dem Phloem der Kormophyten (sie haben sogar Kallosepfropfen in den Siebzellen). Die Transportgeschwindigkeit im Stiel konnte durch Einsatz C-14-markierter Verbindungen bestimmt werden. Bei Macrocystis beträgt sie 70 cm/Stunde, bei Laminaria mit ihren weniger gut entwickelten "Leitbündeln" 5 cm/Stunde.
Die sexuelle Fortpflanzung der Laminariales unterscheidet sich grundsätzlich von der der gleich zu besprechenden Fucales. Der Laminarienthallus ist - wie erwähnt - ein Sporophyt, auf dem sich die Sporangien entwickeln können. Bei den Laminarien unserer Breiten geschieht das ausschließlich zu Beginn des Winters. In den Sporangien erfolgt die Meiose, es werden bewegliche Zoosporen gebildet, die sich, wie bei Ectocarpus, zur Hälfte zu männlichen, zur Hälfte zu weiblichen Gametophyten entwickeln. Diese bestehen aus mikroskopisch kleinen verzweigten Zellfäden. Ihr Wachstum und die Ausbildung reifer Gameten wird durch die Temperatur gesteuert. Bei hoher Temperatur erfolgt rasches Wachstum (zahlreiche Zellteilungen), doch die Reifung der Gameten unterbleibt, bei niederer Temperatur (4-10 Grad C) kommt es rasch zur Bildung reifer Gameten: Spermatozoiden und Eizellen. Als zweiter steuernder Faktor wirkt das Licht. Bei Laminaria saccharina unterbleibt die Gametenbildung in dunklen Wintermonaten und wird erst nach Überschreiten einer kritischen Tageslänge induziert (K. LÜNING und M. J. DRING, Biologische Anstalt Helgoland, 1972).

Die Fucales sind eine sehr große Ordnung, mit zahlreichen, morphologisch deutlich voneinander unterscheidbaren Arten. Die Thalli der Gattung Fucus sind bandförmig gebaut und vielfach verzweigt. Die Bänder sind durch verdickte Zentralstränge verstärkt. Viele Arten besitzen Schwimmblasen; sessile Arten verfügen über Haftkrallen oder eine Haftscheibe. Bei Arten anderer Gattungen ist (z.B. Sargassum linifolium s. nebenstehendes Bild) ist der Thallus noch weiter untergliedert, er ähnelt dem Kormus mancher Landpflanzen, zur Steigerung des Auftriebs ist er mit zahlreichen Schwimmblasen ausgestattet. Er besteht aus einem verzweigten, blattähnliche Thallusanteile tragenden Stiel.
Das Wachstum der Fucales erfolgt durch Teilungen einer Scheitelzelle, die in einer Grube an den Enden wachsender Thallusabschnitte liegt. Im Gegensatz zur Scheitelzelle von Dictyota kann sie Zellen nach verschiedenen Richtungen hin abgeben. Diese bleiben zunächst teilungsfähig (meristematisch), so daß hier durchaus Verhältnisse vorliegen, wie wir sie von den Vegetationspunkten höherer Pflanzen her kennen. Je nach Richtung, in die die Zellen abgesondert werden, differenzieren sie sich zu gewebebildenden Rinden- und Markzellen (Cortex und Medulla).
Zu den in der gemäßigten Zone der nördlichen Hemisphäre häufigsten Arten zählen Fucus vesiculosus (Blasentang) und Fucus serratus (Sägetang; erkennbar an gesägten Thallusrändern). Sie sind die auffallendsten Tange der deutschen Nord- und Ostseeküste. Eine weitere Gattung wäre Himanthalia (Riementang) mit einem deutlich dichotom strukturierten Thallus.
Arten der Gattung Sargassum (Sargassum natans, Sargassum fluitans und ca. 150 weitere) leben, wie schon vermerkt, pelagisch. Sie vermehren sich ausschließlich vegetativ. Die getrenntgeschlechtlichen Gametophyten sind diploid; die Meiose erfolgt in terminal angelegten Rezeptakeln. Das sind angeschwollene Thallusabschnitte mit eingesenkten Konzeptakeln, die ihrerseits Antheridien, respektive Oogonien enthalten. Die Befruchtung der Eizellen erfolgt außerhalb der Gametophyten, die Freisetzung der Gameten unterliegt wieder einer lunaren Rhythmik, die Anlockung der Spermatozoide geschieht chemotaktisch. Die eine Hälfte der Zygoten entwickelt sich zu männlichen, die andere zu weiblichen Gametophyten.
Fucus vesiculosus und Fucus serratus sind ein sympatrisches Artenpaar . Bei Fucus vesiculosus erfolgt die Gametenbildung im wesentlichen im Frühjahr und im Sommer, bei Fucus serratus im Herbst und Winter; somit bleiben sie saisonal voneinander isoliert (M. KNIGHT und M. PARKER, 1950).


Die vier vorgestellten Fälle sollten die Vielgestaltigkeit der Thallusmorphologie und der Fortpflanzungsverhältnisse der Phaeophyta unterstreichen. Dadurch sollte deutlich werden, daß im Verlauf der Evolution der einzelnen Entwicklungslinien unterschiedliche Strategien verfolgt wurden und daß es daher nicht möglich ist, einen für alle Arten gemeinsamen Bauplan anzugeben. Die Thalli der Fucales und der Laminariales sind stets diploid, doch sind die Ursachen der Diploidie verschieden. Die beschriebenen Beispiele sind als Modelle anzusehen, zwischen denen es zahlreiche Übergänge gibt.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de