
This relationship may be illustrated by joint work with the Royal Botanic Gardens, Kew, which examined species richness for five plant families (Dichapetalaceae, Lecythidaceae, Caryocaraceae, Chrysobalanaceae and Proteaceae genus Panopsis) from the long-running Flora Neotropica project among one-degree grid cells (ref 5). The five families include 729 species, and the best available estimate of character richness had to be derived from the current taxonomic classification. Although this is likely to underestimate the degree of character divergence between some species, this is the same method that is used for the bumble bee example in the description of estimates of genetic or character richness, and yet the relationship between species richness and character richness for this much larger group is much closer (below):
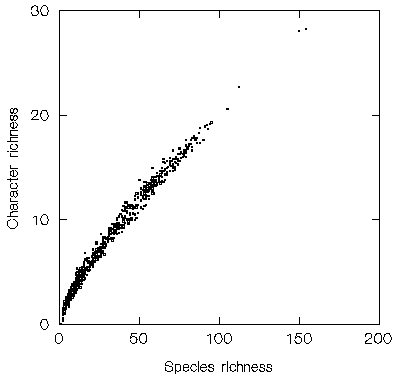
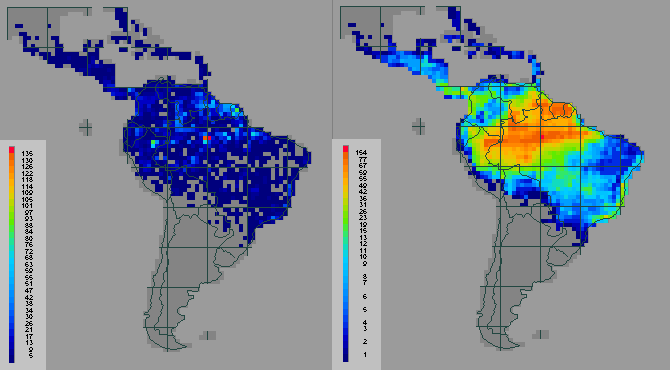
However, using species richness as a surrogate for gene or character richness is still not a panacea, because there are probably too many species even within a suburban garden for complete enumeration, let alone for surveys extending across large regions, such as South America. The problems are illustrated by the same Flora Neotropica data. The five families include 729 species among 1751 grid cells, thereby requiring over one million presence/absence records to be established. Such an enormous sampling effort cannot be deployed quickly and is very expensive. The example shows relative counts of the numbers of species from these five plant families (before, left, and after, right, interpolating the sample data to model the expected distributions of species), with the richer areas shown in orange and the poorer areas shown in dark blue (the score classes on the left are arranged in equal richness intervals, whereas on the right they are arranged for equal numbers of grid cells) (ref 5) (below):
The use of surrogates, in the broadest sense, may be applied to address two problems: first, just as in the Flora Neotropica example for five families of trees above, how good is the species richness of tree floras at predicting the character richness of tree floras?; and second, how good is it for predicting the species and character richness of other groups, or of entire biotas, as an 'indicator group'? The latter, inter-group relationship can be predictive under some circumstances. However, indicator relationships cannot always be assumed, because they can also be weak, absent or even negative, perhaps particularly when indicator and target organisms differ in their habitat associations because different factors govern their distributions.
The geographical patterns of diversity for two groups of organisms can be compared graphically by overlaying the two maps in two separate colours (ref 14). Here, increasing intensity of green is used to represent increasing species richness of butterflies in the first map (data used with permission of the Biological Records Centre), and increasing intensity of blue is used for species richness of birds in the second map (data used with permission of the British Trust for Ornithology). These green and blue maps are then overlaid in the third map. Consequently, black grid cells on the third map show low richness for both butterflies and birds; white shows high richness for both; and shades of grey show intermediate and covarying richness for both (these covarying scores lie on the diagonal of the colour key, to the left of the third map). In contrast, areas of the third map with highly saturated green cells show an excess of richness for butterflies over birds, and areas with highly saturated blue show an excess of birds over butterflies (Spearman correlation coefficient rho= 0.25). The colour classes are arranged to give even frequency distributions of richness scores along both axes (at least within the constraints imposed by tied richness scores) (below):
 |
Link to image building up overlays of butterflies and birds. |
|
|
Link to images showing overlay comparison of birds, butterflies and dragonflies. |
|
|
Link to images showing overlay comparison of Pinaceae and Fagaceae. |