
| der Geißel- und Cilienbewegung, | |
| der Bewegung der Chromosomen während der Mitose und der Meiose und | |
| dem Transport von Granula und Vesikeln in den Zellen und damit verbunden mit der Bildung der Zellwand und der Form und Spezialisierung der Zellen. |
Sie können zu komplexen, regelmäßigen Strukturen vereint sein. Alle Geißeln und Cilien der Eukaryoten zeichnen sich durch den sogenannten "9+2"-Aufbau aus.
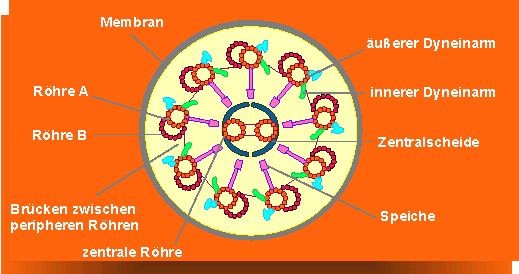
In Pflanzenzellen liegen die Mikrotubuli im Interphasestadium dicht unterhalb des Plasmalemmas (corticale Lage), und ihre Orientierung ist mit der der Cellulosefibrillen in der Zellwand eng korreliert. 1963 war ein entscheidendes Jahr in der Erforschung ihrer Struktur, D. B. SLAUTTERBACK sowie M. C. LEDBETTER und K. R. PORTER (Rockefeller University, New York) setzten erstmals Glutaraldehyd zur Fixierung elektronenmikroskopischer Präparate ein und erreichten damit eine Stabilisierung der Mikrotubulistruktur sowie eine leichtere Darstellbarkeit. Im elektronenmikroskopischen Bild erscheinen sie als charakteristisch gebaute Röhren mit einem äußeren Durchmesser von 240 Å und einem inneren von 140 Å, folglich beträgt die Wandstärke 50 Å. Mikrotubuli bestehen, wie schon beschrieben, aus Tubulin, das in helicaler Anordnung polymerisiert. Zur Ausbildung einer Drehung werden 13 Tubulindimere benötigt. In Geißeln unterscheidet man zwischen einer vollständigen A- und einer unvollständigen B-Röhre, welche sich durch die Art der lateralen Bindungen zwischen den Tubulindimeren (in A: alpha - beta, beta - alpha, in B: alpha - alpha, beta - beta), unterscheiden.
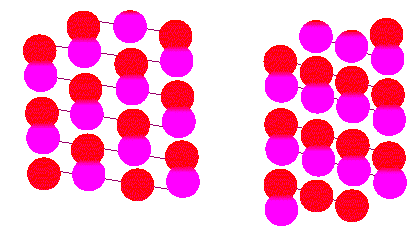
Mikrotubuli sind Röhren aus polymerisiertem Tubulin. Tubulin ist ein Proteindimeres, es besteht aus den beiden Untereinheiten alpha und beta. In der Abbildung sind die Röhren (oder Zylinder) aufgeschnitten dargestellt, so daß die Oberflächenstruktur in Aufsicht erscheint. Die beiden Darstellungen unterscheiden sich durch die Art der Kontakte zwischen den Tubulindimeren (Nach R. E. STEPHENS , K. T. EDDS, 1976).
Wie mit den Mikrofilamenten, so sind auch mit den Mikrotubuli zahlreiche Proteine assoziiert. Das bekannteste ist das Dynein (eine ATPase), das zur Energieumsetzung in Bewegung benötigt wird. Ferner gibt es eine ganze Gruppe von Proteinen mit dem Sammelnamen MAPs (microtubule associated proteins); die meisten von ihnen nehmen offenbar regulatorische Funktionen wahr.
Im Cytoplasma der Zellen liegt in der Regel ein Gleichgewicht zwischen polymerisiertem und depolymerisiertem Tubulin vor, in den Geißeln findet man es nur in polymerisierter Form. Colchicin (das Alkaloid der Herbstzeitlosen Colchicum autumnale), niedrige Temperatur, und ein Überschuß an Calcium-Ionen fördern eine Depolymerisation. Das Alkaloid des Immergrüns (Vinca rosea), das Vinblastin), fällt Tubulin aus.
Es sieht so aus, als würde eine jede Zelle über Kontrollmechanismen verfügen, die die Initiation der Mikrotubulibildung, deren Polymerisation und Depolymerisation, sowie ihre Orientierung in der Zelle regeln. Im Gegensatz zu den Mikrofilamenten gibt es genügend Untersuchungen über ihre Bedeutung in Pflanzenzellen. Antikörper, die gegen tierisches Tubulin gerichtet sind, reagieren auch mit pflanzlichem. Die hohe serologische Verwandtschaft weist darauf hin, daß sie sich im Verlauf der Eukaryotenevolution nur wenig verändert haben und daß ihre Bedeutung für die Zelle bereits frühzeitig festgeschrieben wurde. Der Einfachheit halber soll zunächst ihre Mitwirkung an der Geißel- (und Cilien-) Bewegung, anschließend ihre Bedeutung für Veränderungen im Verlauf des Zellzyklus besprochen werden.
Geißel- und Cilienbewegungen der Eukaryoten (!) sind genaugenommen intrazelluläre Bewegungen; denn obwohl eine Geißel scheinbar als Fortsatz aus einer Zelle herausragt, bleibt sie von der Plasmamembran umschlossen. Alle Dyneinmoleküle entlang der gesamten Mikrotubulilänge müssen kontinuierlich mit ausreichenden ATP-Mengen versorgt werden, das Ionenmilieu und der pH-Wert in ihrer Umgebung müssen stimmen.
Den Bewegungsmechanismus beschrieb P. SATIR (1968, 1976, seinerzeit University of California, Berkeley) als einen sliding filament mechanism, bei dem sich die peripheren Tubulindoubletts aneinander vorbeischieben, wobei das Dynein, das stets an der A-Röhre verankert ist, mit seinen Spitzen Kontakte zur B-Röhre des benachbarten Doubletts schließt.
Der Mechanismus erklärt das Vor- und Zurückschlagen der Geißel, nicht jedoch die zahlreichen Varianten der Geißelbewegungen, die - meist an Einzellern - festgestellt wurden. Es gibt nämlich Zug- und Schubgeißeln sowie Flimmergeißeln. Es gibt solche, die um eine imaginäre Achse rotieren, und es gibt flexible Geißeln, bei denen sich die Bewegung wellenförmig entlang ihrer Achse ausbreitet. Viele Zellen können vom Vorwärts- in den Rückwärtsgang schalten oder mehr oder weniger starke Kurskorrekturen vornehmen.
Cilien unterscheiden sich nur durch ihre Zahl pro Zelle von Geißeln, meist sind sie zudem recht kurz, und oft ist der gesamte Zellkörper von ihnen umgeben. Bei Pflanzen sind sie nur selten anzutreffen, ein immer wieder zitiertes Beispiel sind die Zoosporen von Vaucheria sessilis.
In der Gruppe der Algen (mit Ausnahme der Rotalgen) sind geißeltragende Stadien verbreitet. Ebenso findet man sie bei den Spermatozoiden (männliche Gameten) der Moose und Farne. Im Verlauf der frühen Evolution der Samenpflanzen sind geißeltragende Stadien mehr und mehr zurückgedrängt worden. Zu den wenigen heute noch vorkommenden Ausnahmen gehören die Spermatozoiden von Ginkgo biloba und der Cycadeen.
Oft wird die Bewegung durch externe Signale gesteuert. Viele Einzeller (Algen, Protisten) bewegen sich auf bestimmte Reizquellen zu (Taxien): Licht (phototaktisches Verhalten), bestimmte Chemikalien (chemotaktisches Verhalten). Meist folgt die Bewegung einem Konzentrations- oder Intensitätsgradienten. Wird eine Reizschwelle überschritten (z. B. zu hohe Lichtintensität) setzt eine Umkehrreaktion ein. In den letzten Jahrzehnten wurde viel über die Signalerkennung gearbeitet. So wissen wir z. B., daß die Carotinoide im Augenfleck mancher Algen (z.B. Euglena) blaulichtempfindlich sind, die Chloroplastenbewegung der Alge Mougeotia unter Kontrolle des Phytochromsystems steht und Gameten (der Algen) auf artspezifische Sexuallockstoffe reagieren. Wie die Umsetzung des wahrgenommenen Signals und die Koordination gleich- oder entgegengesetzt gerichteter Signale in eine gerichtete Bewegung aussieht, ist jedoch nicht einmal in Ansätzen bekannt (black box).
An der Basis vieler Geißeln sind oft komplex strukturierte Basalkörper mit Geißelwurzeln vorhanden. M. MELKONIAN (Botanisches Institut der Universität zu Köln) hat sie bei einer Anzahl von Algengruppen analysiert und gruppenspezifische Muster festgestellt. Er wertet diese Strukturen und deren Abänderungen als Merkmale, die wesentlich zum Verständnis der verwandtschaftlichen Beziehungen der einzelnen Algengruppen untereinander beitragen können.
Man weiß seit langem, daß Mikrotubuli den Hauptanteil der mitotischen Kernspindel ausmachen und daß sie die Chromosomenbewegung verursachen. Unklar blieb lange Zeit die Frage nach ihrem Verbleib während der Interphase. Die bereits erwähnte Glutaraldehydfixierung erlaubte es, eine Vielzahl elektronenmikroskopischer Präparate durchzumustern. Immer wieder zeigte es sich dabei, daß die Mikrotubuli vornehmlich an der Plasmaperipherie zu finden sind. In vielen Fällen fand man, daß ihre Anordnung mit der Anlagerung von Cellulosemikrofibrillen bei der sich bildenden Primärwand einhergeht (J. D. PICKETT-HEAPS und D. H. NORTHCOTE, 1966). Damit lag die Annahme nahe, daß ihre Orientierung die Ausrichtung der Mikrofibrillen in der Wand determiniert. Obwohl es eine Menge von Einwänden hiergegen gab, scheint sich die Annahme wohl doch zu bewahrheiten. Es konnte aber auch sichergestellt werden, daß die Existenz der Mikrotubuli nicht Ursache und Voraussetzung einer Zellwandbildung sind, denn in colchicinbehandelten Zellen wird eine Wand gebildet; die Mikrofibrillen sind dann jedoch nicht geordnet, sondern werden in statistischer Anordnung angelagert. Man vermutete zunächst, daß die Mikrotubuli lediglich am Transport von Material beteiligt seien, welches für die Cellulosesynthese benötigt wird. Ebensogut wäre vorstellbar, daß sie die Cellulosesynthetasemoleküle in der Plasmamembran ausrichten, so daß als Folge davon das spezifische Cellulosefibrillen-Muster entsteht (B. A. PALEVITZ und P. K. HEPLER, 1976). Beobachtungen an einigen Algen und an Wurzelhaaren weisen deutlich auf einen solchen Zusammenhang hin. Mikrotubuli können im Plasma untereinander, mit dem Plasmalemma und/oder der Kernhülle in Verbindung stehen.
In Plasmaschläuchen wirken Mikrotubuli an der Verschiebung von sekretorischen Vesikeln, Organellen und anderen zellulären Partikeln mit. Besonders eindrucksvoll ist ein enges Netzwerk, das den wandernden generativen Kern umgibt. Die Spitze des Pollenschlauchs (Apex) enthält keine Mikrotubuli, statt dessen findet man dort Mikrofilamente in erhöhter Zahl.
Eine Voraussetzung der Teilung isolierter Protoplasten ist die Ausbildung einer Zellwand. Ihrer Bildung geht die Umorganisation des peripheren Mikrotubulisystems voraus, wobei die Anordnung der Mikrotubuli die Form der sich bildenden Zelle determiniert. Während die bisher beschriebenen Eigenschaften allgemeiner Natur sind, fallen den Mikrotubuli in spezialisierten Zellen zusätzliche Aufgaben zu. So wirken sie an der Steuerung der Organisation von Zellwandverdickungen und bei der Lignineinlagerung in Sekundärwände mit. Nach Colchicinbehandlung erscheinen nicht die sonst üblichen sekundären Verstärkungsleisten, sondern es bildet sich, analog der Erscheinung bei der Primärwandbildung, ein ungeordnetes Muster an Verstärkungselementen (P. K. HEPLER und E. H. NEWCOMB, 1964, J. CRONSHAW, 1967).
In Zellkulturen konnten M. M. FALCONER und R. W. SEAGULL (1985) die Bildung von Tracheiden analysieren. Fluoreszenzmikroskopische Untersuchungen zeigten deutlich, daß Mikrotubuli überall dort lokalisiert sind, wo es zur Ausbildung von tracheidalen Wandverstärkungen kommt. Das Muster dieser Wandverstärkungen stimmte mit dem Muster der Mikrotubulianordnung überein.
Ungleiche Wanddicken und eine Formveränderung der Zelle, so wie wir sie z. B. bei der Bildung von Stomatazellen vorfinden, beruhen womöglich auch auf ungleicher Verteilung intrazellulärer Mikrotubuli während der Morphogenese. Sie sind an den Seiten mit verdickten Wänden (den periklinen Wänden) häufiger als an jenen mit dünnen (den antiklinen Wänden).
Die Menge an Mikrotubuli ist mit einer Anhäufung von Dictyosomen korreliert, so daß es so aussieht, als würden sie tatsächlich für die Ausrichtung des Materialtransports und die Verteilung des Zellwandmaterials in der Zelle benötigt (B. A. PALEVITZ und P. K. HEPLER, 1976, B. GALATIS, 1980). In manchen Algen (z. B. Hydrodictyon, Pediastrum, Staurastrum) beeinflussen sie die Zellform und die Koloniebildung (H. J. MARCHANT und J. PICKETT-HEAPS, 1974, H. J. MARCHANT, 1979) und bei Micrasterias die postmitotische Kernwanderung (O. KIERMEYER, 1972). Hier haben sie keinen Einfluß auf die Formbildung der Zellen. Bei dem zellwandlosen Flagellaten Ochromonas hingegen ist die Anordnung der Mikrotubuli für die Ausbildung der spezifischen Zellform verantwortlich; bei Euglena ist eine entsprechende Anordnung in der Pellikula erkennbar. Dort wirken sie an den ständigen Formveränderungen der Zellen ("metabolische Bewegung") mit.
Wie schon erwähnt, wirken Mikrotubuli, wie auch die vorher besprochenen Mikrofilamente an der intrazellulären Plasmabewegung und somit auch an der Bewegung der Organellen und anderer Partikel mit. Damit stellt sich die Frage, welches der beiden Systeme für bestimmte Bewegungsabläufe primär verantwortlich sei. Die Antwort darauf ist artspezifisch. Bei siphonalen Grünalgen fand D. MENZEL (Universität Heidelberg, 1987), daß die Bewegung der Organellen in der Gattung Caulerpa durch Mikrotubuli erfolgt, bei Acetabularia durch Mikrofilamente und daß bei Bryopsis beide Systeme gleichermaßen benötigt werden.
 Was geschieht, wenn in einer Pflanzenzelle die Vorbereitungen zur Mitose
(oder Meiose) einsetzen? Ein besonders auffälliges Merkmal vieler
solcher Zellen ist die Ausbildung eines Präprophasebandes, das den
Kern umgibt und in der Ebene des später zu bildenden Phragmoplasten
liegt. Es entsteht durch Verlagerung nahezu des gesamten Tubulins der Zelle
in diese Region, d.h., das tubuläre Cytoskelett des Interphasestadiums
wird abgebaut, ein neues wird errichtet (P. K. HEPLER und D. H. NORTHCOTE,
1967). Es gibt Zellen, z.B. die vielstudierten Endospermzellen von Haemanthus
katherinae, denen das Band fehlt. Ob es erforderlich ist, um den Zellkern
in die richtige (zentrale) Position zu bringen, oder ob beide Prozesse
unabhängig voneinander ablaufen, war eine lange umstrittene Frage.
Obwohl noch nichts endgültig entschieden ist, tendiert man heutzutage
zu der Annahme, daß beide Ereignisse voneinander unabhängig
seien. Es ist aber auch denkbar, daß die Lage des Bandes die Orientierung
der Teilungs- oder Mitosespindel determiniert. Mit dem Einsetzen der Prophase
beginnt ihre Errichtung. Es besteht allgemein Konsens darüber, daß
sie aus Mikrotubuli besteht, doch weit schwerer ist die Frage zu beantworten,
wie sie die Chromosomenbewegung bewirken. Zunächst: Es gibt zwei Organisationstypen,
zu denen sich Mikrotubuli zusammenschließen: Mikrotubuli, die von
den Polen ausgehend bis weit über die Äquatorialebene hinausreichen
können, nennt man polare Mikrotubuli. Sie überlappen einander,
so daß es so aussieht, als ob sie von Pol zu Pol reichten. Andere
Mikrotubuli setzen an den Kinetochoren (= Centromeren) der Chromosomen
an und reichen bis in die Nähe der Pole oder sind mit den polaren
Mikrotubuli assoziiert. Man nennt sie chromosomale oder Kinetochor-Mikrotubuli.
Was geschieht, wenn in einer Pflanzenzelle die Vorbereitungen zur Mitose
(oder Meiose) einsetzen? Ein besonders auffälliges Merkmal vieler
solcher Zellen ist die Ausbildung eines Präprophasebandes, das den
Kern umgibt und in der Ebene des später zu bildenden Phragmoplasten
liegt. Es entsteht durch Verlagerung nahezu des gesamten Tubulins der Zelle
in diese Region, d.h., das tubuläre Cytoskelett des Interphasestadiums
wird abgebaut, ein neues wird errichtet (P. K. HEPLER und D. H. NORTHCOTE,
1967). Es gibt Zellen, z.B. die vielstudierten Endospermzellen von Haemanthus
katherinae, denen das Band fehlt. Ob es erforderlich ist, um den Zellkern
in die richtige (zentrale) Position zu bringen, oder ob beide Prozesse
unabhängig voneinander ablaufen, war eine lange umstrittene Frage.
Obwohl noch nichts endgültig entschieden ist, tendiert man heutzutage
zu der Annahme, daß beide Ereignisse voneinander unabhängig
seien. Es ist aber auch denkbar, daß die Lage des Bandes die Orientierung
der Teilungs- oder Mitosespindel determiniert. Mit dem Einsetzen der Prophase
beginnt ihre Errichtung. Es besteht allgemein Konsens darüber, daß
sie aus Mikrotubuli besteht, doch weit schwerer ist die Frage zu beantworten,
wie sie die Chromosomenbewegung bewirken. Zunächst: Es gibt zwei Organisationstypen,
zu denen sich Mikrotubuli zusammenschließen: Mikrotubuli, die von
den Polen ausgehend bis weit über die Äquatorialebene hinausreichen
können, nennt man polare Mikrotubuli. Sie überlappen einander,
so daß es so aussieht, als ob sie von Pol zu Pol reichten. Andere
Mikrotubuli setzen an den Kinetochoren (= Centromeren) der Chromosomen
an und reichen bis in die Nähe der Pole oder sind mit den polaren
Mikrotubuli assoziiert. Man nennt sie chromosomale oder Kinetochor-Mikrotubuli.
Erstere entstehen durch ein an den Polen beginnendes Wachstum, letztere beginnen mit dem Wachstum (der Initiation des Polymerisationsprozesses) an den Centromeren (Kinetochoren).
In tierischen Zellen sind an den Polen Centriolen erkennbar. In ihrer Nähe liegen die Kondensationspunkte, von denen die Mikrotubuli ausgehen. Den meisten - vor allem den höheren - Pflanzen fehlen sie. Bei einigen Algen kommen sie vor, bei den Characeen treten sie in der Meiose, nicht jedoch in der Mitose in Erscheinung. Sie sind demnach, zumindest bei Pflanzen, für die Bildung der polaren Mikrotubuli entbehrlich. Was an ihrer Stelle steht, ist hingegen nicht bekannt.
Wie aus zahlreichen cytologischen Befunden hervorgeht, können nur kinetochorenhaltige Chromosomen bewegt werden. Das heißt, daß es eine spezifische Bindungsstelle für Mikrotubuli am Chromosom gibt (sie kann bei Chromosomen mit "diffusem" Centromer die gesamte Chromosomenoberfläche einschließen). Die Wanderung zu den Polen beruht auf einer Interaktion zwischen den polaren und den Kinetochor-Mikrotubuli. Aller Voraussicht nach werden die Chromosomen gezogen, nicht gestoßen. Die Bewegung beruht sicher nicht auf einem einfachen Gleitmechanismus. Die Chromosomenbewegung in der Anaphase kommt vielmehr durch Verkürzung der Kinetochor-Mikrotubuli am Kinetochorpol, bei gleichzeitiger Verlängerung der polaren Mikrotubuli zustande (McINTOSH und KOONCE, 1989).
Offen bleibt die Frage nach der Energieübertragung. Während der Telophase löst sich die Kernspindel auf, die Mikrotubuli sammeln sich im Bereich des Phragmoplasten, von wo aus die Organisation des Interphase-Cytoskeletts de novo beginnt. Die Mikrotubuli liegen in der Phragmoplastenebene in engster Nachbarschaft zu den Mikrofilamenten. Wichtig wäre daher die Frage nach der Kooperation bzw. der Arbeitsteilung zwischen beiden Systemen, doch auch hierzu gibt es noch nichts zu sagen.
|
|