 |
Normalerweise sind Zell- und Kernteilung miteinander gekoppelt. Mit Ausnahmen werden wir uns an anderer Stelle befassen. Der Ablauf der Kernteilung in pflanzlichen Zellen wurde von E. STRASBURGER aufgeklärt, 1875 erschien sein Standardwerk "Zellbildung und Zellteilung". Schon 1884 waren die Befunde bereits Inhalt seines "Kleinen botanischen Praktikums", und seit 1894 sind sie ein fest etablierter Bestandteil des "Lehrbuchs der Botanik". In der 4. Auflage (1900) wird die Kernteilung wie folgt beschrieben:
"Von bestimmten, ganz begrenzten Fällen abgesehen, vermehren sich die pflanzlichen Zellkerne auf dem Wege der sog. mitotischen oder indirekten Teilung. Der Teilungsvorgang wird auch Karyokinese (Anm.: heute üblicherweise Mitose) bezeichnet. Er spielt sich in ziemlich komplizierter Weise ab, die aber notwendig erscheint, um die Substanz des Mutterkerns völlig gleichmäßig auf die beiden Tochterkerne zu verteilen."
Anfangs arbeitete STRASBURGER mit alkoholfixiertem Material. 1879 konnte A. LUNDSTRÖM die Mitose am lebenden Objekt (Staubfadenhaaren von Tradescantia) untersuchen. Mit modernen mikroskopischen Techniken, vor allem der Phasenkontrast- und der Interferenzkontrastmikroskopie, lassen sich heutzutage die Vorgänge der Kernteilung an einer Vielzahl von Zelltypen sichtbar machen.
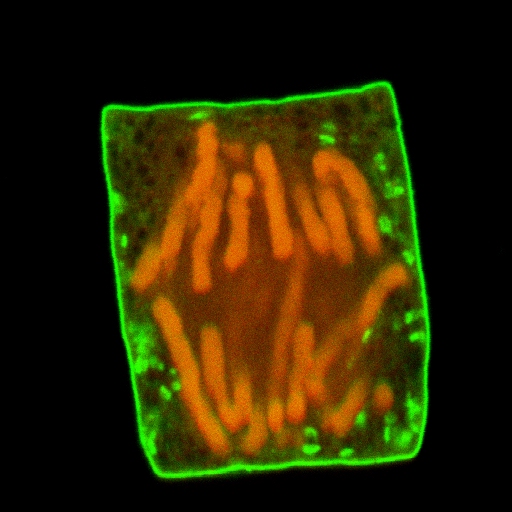
Zwiebelzelle aus der Wurzelspitze, aufgenommen mit Konfokalem Laser Scanning Mikroskop: Anaphae, Chromosomen rot fluoreszierend - Plasmamembran und Golgi Apparat grün - |
Das Stadium des Sichtbarwerdens der Chromosomen zu Beginn der Mitose wird auf STRASBURGERs Vorschlag hin Prophase genannt. Der Kieler Anatom W. FLEMMING erkannte 1880, daß sich Chromosomen der Prophase durch einen Längsspalt auszeichnen.
Im Anschluß an die Prophase ordnen sich die Chromosomen in der Äquatorialebene (Zellteilungsebene) an und bilden somit die Äquatorialplatte. Dieses Stadium ist die Metaphase. Die beiden Chromosomenlängshälften (=Chromatiden) treten dabei deutlich in Erscheinung. Sie rücken hierauf in entgegengesetzte Richtungen auseinander, um die beiden Tochterkerne zu bilden. Die Phase des Auseinanderweichens wird Anaphase, die der Bildung von Tochterkernen Telophase genannt.

In die geschilderten Vorgänge greifen andere wiederum in
bestimmter Weise ein. Während die Kernfäden kürzer
werden, sich entwirren und in die einzelnen Chromosomen zerfallen,
bilden sich im Plasma faserig aussehende Aggregate, die sich in
der Zelle von Pol zu Pol anordnen. Man nennt sie Spindelfasern
und die gesamte Struktur Kernspindel. Die Spindel besteht
aus "Fasern" (das sind Aggregate oder Bündel, bestehend aus sich überlappenden Mikrotubuli, die ihrerseits kürzer als das Bündel selbst sind), die von Pol zu Pol reichen, und solchen,
die einen Pol mit einem Chromosom verbinden (Stützfasern
und Zugfasern). Das Auseinanderweichen der Chromatiden beruht
auf einer Kontraktion der Zugfasern. Eine Zugfaser setzt stets
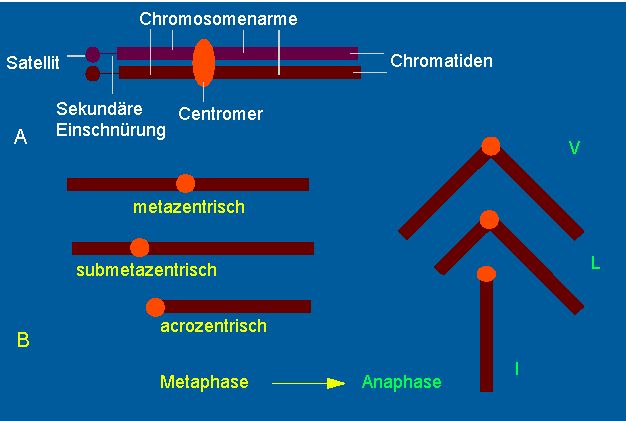
an einer bestimmten Stelle des Chromosoms, dem Centromer
oder der Spindelansatzstelle an. Entsprechend der Lage
des Centromers entstehen die für die Anaphase typischen V-,
L-, U- oder I-förmigen Strukturen .
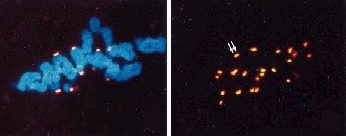
Immunochytochemische Methoden und Einsatz der Konfokalen Laser Scan Mikroskopie erlauben es, die Centromere eindrucksvoll abzubilden. Die Chromosomen - durch Propidium Jodid fluorochromiert - sind im linken Bild erkennbar. Versuchsobjekt Lilium longiflorum. |
Heute weiß man, daß die Fasern Tubulin enthalten. Unter Verwendung spezifischer, fluoreszenzmarkierter Antikörper läßt es sich in der Kernspindel selektiv darstellen. Während der späten Anaphase wird in den Pflanzenzellen in der Ebene der Äquatorialplatte die Anlage einer neuen Zellwand sichtbar; die Wandbildung schreitet während der Telophase fort.
Tierische Zellen hingegen teilen sich durch Einschnürung der Mutterzelle. Die entscheidende Bedeutung der Mitose liegt in allen Fällen in der qualitativ und quantitativ stets gleichen Verteilung der Erbanlagen, wobei die Längsspaltung den wesentlichen Schritt darstellt. Jede Tochterzelle erhält folglich eine Chromosomenhälfte (Chromatid). Die Zahl der Chromosomen ist in der Regel artspezifisch.
Auch diese Entdeckung geht auf E. STRASBURGER zurück; der Prager Zoologe RABL dehnte die Aussage auch auf tierische Zellen aus.
Abweichungen von dieser Regel beruhen oft auf dem Verkleben einzelner Chromosomen an ihren Enden, wodurch sich die Chromosomenzahl abartig ändert. Solche Änderungen, verbunden mit gleichzeitigen genetischen Veränderungen, erbrachten schließlich den noch ausstehenden endgültigen Beweis für die Richtigkeit der Chromosomentheorie.
Mitosestadien - bitte Geduld - die einzelnen Stadien erscheinen nacheinander. Rechts im Bild der Mitose- oder Zellzyklus. Außer den einzelnen Phasen der Mitose besteht er aus der G1- Phase (gap-Phase), der S-Phase, in der DNS repliziert wird und der G2-Phase, in der die Chromosomen jeweils zwei Chromatiden enthalten. © StR Holger Schickor - Staatliches Eifel-Gymnasium Neuerburg
|