
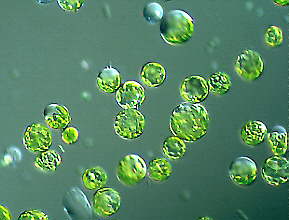
 |
|
- die Mittellamelle durch Pektinasen aufgelöst wird, und dann
- die Zellwand durch Cellulase verdaut wird.
Bei den dazu benötigten Enzymen handelt es sich in der Regel nicht um reine Präparate, sondern um Rohextrakte aus bestimmten Bakterien und Pilzen. Anstelle des Zweistufenverfahrens verkürzt man den Prozeß vielfach auf einen Arbeitsgang, indem man ein Enzymgemisch verabreicht. Wegen des hohen osmotischen Drucks im Protoplasteninneren müssen isotonische Medien verwandt werden. Üblicherweise setzt man ihnen die Zuckeralkohole Mannitol und/oder Sorbitol zu. Ansonsten enthält ein Protoplastenmedium eine Vielzahl anorganischer Salze und einige Vitamine. Protoplasten können darin über Tage am Leben gehalten werden und behalten dabei ihre Teilungsfähigkeit. Sie können auf Agarplatten kultiviert werden, wo sie zu kleinen sichtbaren Kolonien (Kalli) heranwachsen, womit wir wieder bei der "Mikrobiologie" mit ihren vielen Möglichkeiten sind.
Zu den markanten Eigenschaften aktiver Protoplasten gehört die rasche Regeneration einer Zellwand, die einer Protoplastenteilung stets vorangeht. Es sieht ganz so aus, als sei Wandbildung eine ursächliche Voraussetzung für die Zellteilung und die Ausbildung einer Polarität. Bei der Protoplastenherstellung ging diese ebenso verloren wie die Positionsinformation. Beide müssen bei einer Regeneration zu einer Pflanze neu erworben werden.
Protoplasten eignen sich zur Herstellung von "Vakuoplasten" (also reinen Vakuolen) und Subprotoplasten, die lediglich aus Zellkern und einem Plasmasaum bestehen. Die einzelnen Fraktionen können durch Dichtegradientenzentrifugation voneinander getrennt und angereichert werden. Wozu sind Protoplasten zu gebrauchen?
- Protoplasten können zu vollständigen Pflanzen regeneriert werden.
- Protoplasten gleicher oder unterschiedlicher Herkunft können miteinander fusioniert werden. Aus dem Fusionsprodukt kann ggf. eine Pflanze regeneriert werden (somatische Bastardierung).
- Makromoleküle (Nukleinsäuren und Proteine), Viren, Zellbestandteile (Chromosomen, Chloroplasten usw.) können von den Protoplasten durch Phagozytose aufgenommen werden.
- Protoplasten eignen sich zum Studium der molekularen Architektur von Pflanzenzellen.
1970 haben I. TAKEBE, G. LABIB und G. MELCHERS (Max-Planck-Institut für Biologie, Tübingen) aus Protoplasten des Tabaks eine vollständige Pflanzen regeneriert.
Y.Y. GLEBA (Akademie der Wissenschaften der Ukrainischen SSR, Kiew) entwickelte ein Verfahren zur Kultur isolierter Protoplasten in Mikrotropfen. Eine Anzahl derart klonierter Protoplasten konnte zur Entwicklung kompletter Pflanzen gebracht werden.
Es gibt mittlerweile eine Reihe dikotyler Angiospermen, aus deren Protoplasten vollständige fertile Pflanzen regeneriert wurden. Die geringsten Schwierigkeiten bereiten die Solanaceen. Protoplasten aus Leguminosen und Monokotyledonen, hingegen, konnten nur nach Entwicklung aufwendiger Verfahren und auch nicht immer zur Regeneration ganzer Pflanzen gebracht werden. Für eine Regeneration ist ein Zusatz von Phytohormonen zum Medium erforderlich. Es sieht so aus, als würden den Mesophyllzellen der Monokotyledonen die notwendigen Rezeptoren verloren gehen. Das mag auch der Grund dafür sein, daß hormonähnliche Herbizide Dikotyledonen schädigen, das Monokotyledonenwachstum (Getreide) aber nicht beeinträchtigen. Andererseits ist das mangelnde Regenerationsvermögen der Protoplasten wirtschaftlich wichtiger Kulturpflanzen ein großes Handicap, die mit dieser Methode gewonnenen Erfahrungen in die landwirtschaftliche Praxis umzusetzen.
Eine Aggregation zweier oder mehrerer Protoplasten genügt nicht, um eine Fusion einzuleiten. Protoplastenoberflächen sind stark negativ geladen. Im Gegensatz zu tierischen Zellen beruht die Oberlächenladung jedoch nicht auf Sialinsäureresten, sondern sie wird durch Phosphatgruppen bedingt. Intakte Protoplasten stoßen einander deshalb in Suspension ab. Zur Neutralisation der Oberflächenladung eignen sich Mono- und Polykationen. Sehr wirkungsvoll sind Calciumionen oder Polyäthylenglycol, wodurch die Protoplasten vernetzt und zur Fusion gebracht werden können. Eine alternative Lösung ist das Anlegen eines elektrischen Feldes.
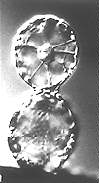
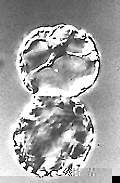
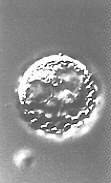
Fusion von Protoplasten im elektrischen Feld (Elektrofusion) (H.-U. KOOP, H.-G. SCHWEIGER, 1985
Bei einer sexuellen Bastardierung fusionieren haploide Zellen, die aus einer vorangegangenen Meiose entstanden sind. Bei der Fusion somatischer diploider Zellen müßte man ein tetraploides Fusionsprodukt erwarten, sofern auch die Zellkerne fusionieren. Wenn das der Fall ist, spricht man von einem Synkaryon; bleiben die Kerne getrennt, von einem Heterokaryon.
Um Verhältnisse zu schaffen, die mit denen einer sexuellen Kreuzung vergleichbar sind, benötigt man haploides Ausgangsmaterial. Zu diesem Zweck entwickelten S. GUHA und S. C. MAHESHWARI (Botanisches Institut der Universität Delhi, 1966) ein Medium zur Kultur der Antheren von Datura innoxia. Aus den Staubbeuteln wuchsen Keimlingen ähnliche Pflänzchen (Embryoide) heran. Dadurch, daß sie nicht die diploide Chromosomenzahl des normalen Pflanzengewebes, sondern nur die haploide hatten, erwiesen sich diese sich nicht zu Pollenkörnern entwickelnden Abkömmlinge einer Meiose als "Pflanzen aus Gonen". Gonen ist die Bezeichnung für die (haploiden) Produkte einer Meiose (unabhängig vom Geschlecht). Es ist mittlerweile gelungen, auf diesem Wege aus einer ganzen Reihe verschiedener Pflanzenarten (Di- und Monokotyledonen) haploide Pflanzen zu erzeugen und sie als Ausgangsmaterial zur Protoplastenherstellung zu nutzen.
Um haploide Pflanzen durch Samen fortpflanzen zu können, müssen sie diploid gemacht werden. Das ist aber, seit 1937 das Colchicin für diese Aufgabe entdeckt wurde, nicht schwer. Man kann sich fragen, worin der Vorteil dieses Verfahrens liegt, und die Antwort lautet: Die so produzierten diploiden Pflanzen sind in allen ihren Genen homozygot.
Haploide Nicotiana-Arten (N. tabacum, N. sylvestris u.a.) sind zunächst für intra-, später auch für interspezifische Fusionen ihrer Protoplasten herangezogen worden. Die durch Regeneration von Fusionsprodukten erzeugten Pflanzen unterscheiden sich nicht von solchen, die auf sexuellem Wege erzeugt wurden. Zur Selektion von Fusionsprodukten eignen sich Elternstämme mit sich gegenseitig komplementierenden Chlorophylldefekten; Fusionsprodukte sind dann an ihrer grünen Färbung erkennbar (G. MELCHERS und G. LABIB, Tübingen 1974).
Interspezifische Fusionen sind relativ leicht herstellbar. Es gelingt sogar, pflanzliche Protoplasten und tierische Zellen (Fibroblasten) zu fusionieren und die Fusionsprodukte einige Stunden am Leben zu erhalten. Es geht bei solchen Versuchen natürlich nicht um Regenerationsversuche, sondern um Fragen der Kooperation der Membranen untereinander oder der Exprimierbarkeit einzelner Gene einer Pflanzenzelle im Tierzellplasma oder umgekehrt. Von wenigen Ausnahmen abgesehen, können interspezifische Fusionsprodukte nur dann regeneriert werden, wenn auch Kreuzungen auf sexuellem Weg erfolgreich sind.
Interspezifische Heterokarien, vor allem bei weniger nah verwandten Arten, bilden keine Synkarien aus, die Kernteilungen sind asynchron, Chromosomen gehen verloren, das System gerät aus dem Gleichgewicht. Dennoch gibt es bereits einige Erfolge interspezifischer somatischer Hybridisierung. G. MELCHERS gelang es 1978, Tomaten- und Kartoffelprotoplasten miteinander zu fusionieren und das Fusionsprodukt (Tomoffel. Karmate) zur Regeneration ganzer Pflanzen zu bringen. Die Pflanzen blühten, fertile Samen wurden aber nicht gebildet.
O. SCHIEDER (Max-Planck-Institut für Züchtungsforschung, Köln) war mit Fusionsprodukten zwischen Datura innoxia und Datura stramonium auch in diesem Punkt erfolgreich. Die aus der Fusion hervorgegangenen Pflanzen sind voll fertil, und daher als neue Art zu betrachten: Datura straubii. Datura-Arten werden zur Gewinnung der medizinisch wichtigen Alkaloide Hyoscyamin und Scopalamin verwendet. Datura straubii ist wüchsiger als die beiden Ausgangsarten, ihr Gehalt an Alkaloiden liegt um 20-25 Prozent über dem der Eltern.
Protoplasten können durch Phagozytose Fremdmoleküle und Organellen aufnehmen. Damit gelang unter anderem der Nachweis, daß eine Zelle durch mehr als nur ein Viruspartikel infizierbar ist und daß sich unterschiedliche Virusstämme in einer Zelle vermehren können. Ferner gelang eine Transformation von Protoplasten mit Ti-DNS aus Agrobacterium tumefaciens, wodurch sich ein Weg eröffnet hat, das System für gentechnische Zwecke und Untersuchungen einzusetzen. Die Aufnahme von Plastiden erlaubt die Analyse der Kooperation zwischen Kern- und Plastidengenom. Die Aufnahme einzelner artfremder Gene kann genutzt werden, um einen genetischen Defekt im Protoplastengenom zu beheben. Somit ließ sich in einer Nitratreduktase-defizienten Mutante von Nicotiana tabacum, nach Fusion mit inaktivierten Protoplasten aus Physalis und Datura, eine Nitratreduktaseaktivität übertragen. Die Transformation erwies sich als stabil (R. P. GUPTA, M. GUPTA, O. SCHIEDER, 1982).
Durch Anwendung moderner analytischer Verfahren (Gelelektrophorese, Nachweis von Enzymaktivitäten im Gel, Isoenzyme usw.) läßt sich der Erfolg von Fusionsexperimenten belegen. Eine wichtige Markerrolle spielt die Ribulose-1,5-Bisphosphatcarboxylase, da sie aus kerncodierten und plastidencodierten Untereinheiten besteht und weil die Analyse dieses Proteins zugleich Auskünfte über Aktivitäten von Plastiden und Kern der Elternarten im Fusionsprodukt gibt.
Nachdem die Protoplastentechniken etabliert waren, war man daran interessiert, Mutationen zu induzieren und die Mutanten zu isolieren; besonderes Interesse galt den auxotrophen Stämmen, also solchen, die auf die Zufuhr bestimmter Stoffe angewiesen sind. Es gibt einige wenige Erfolge in dieser Richtung, doch die Zahl der Mißerfolge ist größer. Ein Grund dafür liegt vermutlich in der Tatsache, daß ein großer Teil der Angiospermengenome allopolyploid ist und daß die genetische Information selbst in Haploiden mehrfach vorliegt, so daß der Defekt in einem Satz durch ein intaktes Gen in einem der anderen kompensiert wird. Hinzu kommt, daß Pflanzen oft auf unterschiedlichen Stoffwechselwegen zum gleichen Produkt gelangen. Ein möglicher Ausweg hieraus wäre die Nutzung von "monohaploiden" Ausgangsformen (1x anstelle von 1n). Im Falle von Kulturpflanzen heißt das, daß man sich zunächst auf die Suche nach ihren Ausgangsformen begeben muß.
Beispiele hierfür sind bereits mehrfach zitiert worden. Da bei der Protoplastenbildung aus nativen Zellen die ursprüngliche Zellform verlorengeht, haben S. M. WICK et al. (Australian National University, Canberra, 1981) Protoplasten aus vorfixierten Zellen hergestellt. Zwar leben sie nicht mehr, haben aber ihre Form behalten, und durch Einsatz der indirekten Immunfluoreszenz läßt sich ein Bild der Verteilung von Strukturelementen in der Zelle gewinnen. Durch Verwendung von Lektinen ließ sich zeigen, daß die Protoplastenoberflächen verschiedener Arten unterschiedliche Kohlenhydratmuster tragen und daß sich die Lektinbindungseigenschaften des Plasmalemmas grundsätzlich von den intrazellulären Membranen und dem Tonoplasten unterscheiden.
|
|