
Spaltöffnungen (Stomata) sind funktionelle Einheiten der Epidermis, die dem Gasaustausch zwischen den Interzellularen des pflanzlichen Gewebes und der Außenwelt dienen. Besonders häufig - und von charakteristischer Gestalt - findet man sie bei der überwiegenden Zahl der Arten in der Epidermis der Blattunterseite. Ihre Entstehungsgeschichte variiert bei den einzelnen Pflanzengruppen, doch sind stets inäquale Teilungen mit im Spiel. Zu einer Funktionseinheit gehören sowohl die eigentlichen, fast immer chloroplastenhaltigen Schließzellen und die benachbarten chloroplastenfreien Nebenzellen.
Es ist allgemein bekannt, daß Spaltöffnungen in feuchter Umgebung geöffnet, in trockener geschlossen sind. J. HEDWIG erkannte im 18. Jahrhundert als erster, daß ihnen die Aufgabe von "Ausdünstungsöffnungen" zufällt. Heute sagen wir dazu, sie seien an der Transpiration beteiligt. Es lag daher auf der Hand, den Mechanismus der Schließbewegung auf Turgoränderungen zurückzuführen und sie als Turgorbewegungen einzustufen. Die pflanzenanatomische Untersuchung der Stomata (vor allem die Betrachtung der Querschnitte) verifizierte diese Annahme. Bei vielen Arten sind die Außen- und Innenwand im Vergleich zu den Seitenwänden stark verdickt. Sie setzen daher jeglicher Deformation einen merklichen Widerstand entgegen.
Die Wand zwischen Schließ- und Nebenzelle und die den Spalt formende Wand sind relativ dünn und daher leicht dehnbar. Diese Asymmetrie im Zellbau und der Wandstärken erklärt die gerichtete, durch Turgor bedingte Bewegung. Die Mechanik der Schließzellen ist damit verständlich. Probleme ergaben sich jedoch bei der Aufklärung der Auslösung und der Regulation dieses Vorgangs: Wasser erwies sich zwar als bedeutender, doch nicht alleiniger Kontrollfaktor der Schließzellbewegung. Als nicht minder wichtige Regelgrößen haben sich
![]() die Kohlendioxyd-Konzentration,
die Kohlendioxyd-Konzentration,
![]() Licht,
Licht,
![]() Kalium-Ionen und
Kalium-Ionen und
![]() Abscisinsäure (ABA) erwiesen.
Abscisinsäure (ABA) erwiesen.
Schließzellen können Wasser nach drei Richtungen abgeben:
- nach außen,
- in die benachbarte Nebenzelle und
- in die Atmungshöhle (einen Teil des Interzellularsystems des Blattgewebes, der unter den Schließzellen liegt).
Bei geöffneten Spalten stellt sich ein Gleichgewicht zwischen den Wasserdampfkonzentrationen der Atmosphäre und der Atemhöhle ein. Solange der Wassernachschub sichergestellt ist, bleiben die Spalten offen. Da es in der Regel ein starkes Wasserpotentialgefälle zwischen feuchtem Boden und meist trockener Atmosphäre gibt, bilden Pflanzen ein zwischengeschaltetes Verteilerelement. Sie ihrerseits profitieren von dem Konzentrationsgefälle (für sie ein Energiegewinn), wobei die Schließbewegungen der Stomata einen entscheidenden regulierenden Einfluß ausüben. Bei zu hohem Wasserverlust bzw. nicht ausreichendem Nachschub schließen sie sich. Bei Spaltenöffnung ist der osmotische Druck in den Schließzellen weit höher als in den Nebenzellen. Bei geschlossenen Spalten verschiebt sich das Verhältnis zugunsten der Nebenzellen.
Bei den meisten Pflanzenarten sind die Spaltöffnungen bei Dunkelheit geschlossen. Licht fördert das Öffnen. Das Aktionsspektrum ähnelt dem der Photosynthese. Blaulicht ist besonders wirksam. In CAM-Pflanzen, z.B. den Crassulaceen, sind die Spaltöffnungen nachts offen. Sie sind nämlich darauf angewiesen, nachts Kohlendioxyd zu akkumulieren, in Form von Malat oder Aspartat zu speichern, um es dann tagsüber in den CALVIN-Zyklus einzuschleusen. Offene Spaltöffnungen am Tage würden an Standorten, die von CAM-Pflanzen besiedelt werden, zu intolerablen Transpirationsverlusten führen.
Eine niedrige Kohlendioxyd-Konzentration (in der Atemhöhle) führt zu einer Spaltenöffnung, eine hohe zu Spaltenverschluß. Bei Dunkelheit entsteht in pflanzlichen Geweben durch Atmung reichlich Kohlendioxyd, so daß die Spalten geschlossen bleiben. Mit dem ersten Tageslicht kann die Photosynthese unverzüglich einsetzen, weil ausreichend Kohlendioxyd akkumuliert worden ist. Da die Schließzellen im Unterschied zu den Nebenzellen Chloroplasten enthalten, findet in ihnen auch eine Photosynthese statt, und diese Aktivität wiederum steht im Zusammenhang mit dem Anstieg des osmotischen Werts und damit der Öffnung der Spalte.
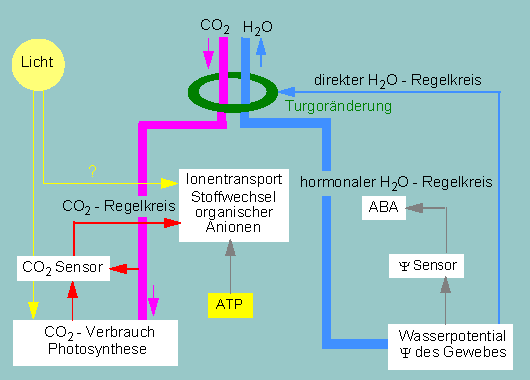
Regelkreise zur Kontrolle der Spaltöffnungsbewegungen. Eine
Schließzelle ist oben schematisch dargestellt.
(Nach K.
RASCHKE, 1975)
Soweit die Beobachtungen und Messungen. Worauf beruhen nun aber die Erhöhung und Erniedrigung des osmotischen Werts? Es gibt gute Hinweise darauf, daß das Licht in erster Linie über die Erniedrigung der inter- und intrazellulären Kohlendioxyd-Konzentration wirkt. Durch die Photosynthese in den Mesophyllzellen wird Kohlendioxyd schneller verbraucht, als es nachgeliefert werden kann. Dadurch entsteht ein interzelluläres Defizit in Spaltöffnungsnähe. Die Photosyntheseaktivität in den Schließzellen selbst sorgt für den Abfall des intrazellulären Kohlendioxyd-Spiegels und eine gleichzeitige Steigerung der Wasseraufnahme aus den Nachbarzellen (Nebenzellen).
Es hat sich gezeigt, daß der Wasseraufnahme eine Kalium-Aufnahme vorangeht. Kalium-Ionen werden durch aktiven Transport (eine Kalium-Pumpe) aus den Nebenzellen in die Vakuolen der Schließzellen verlagert. Parallel dazu erfolgt eine Akkumulation von Anionen (Chlorid, Malat) in den Vakuolen. Im Gegenzug werden Protonen an die Nebenzellen abgegeben. Quantitativ sind die Ionenflüsse ausreichend, um eine für die Schließbewegung erforderliche Turgorzunahme zu erklären, doch ist damit immer noch nichts über den primären Auslöser gesagt. Folgende Fragen wären zu beantworten:
![]() Wodurch kommt die Steigerung der Aktivität der Kalium-Pumpe
zustande?
Wodurch kommt die Steigerung der Aktivität der Kalium-Pumpe
zustande?
![]() Ist der Protonenfluß eine Ursache oder die Folge des
Kalium-Ionentransports?
Ist der Protonenfluß eine Ursache oder die Folge des
Kalium-Ionentransports?
Wie wichtig die Kalium-Pumpe für die Schließzellenbewegung tatsächlich ist, geht aus der Wirkung eines Pilzgifts, des Welketoxins Fusicoccin (aus Fusicoccum amygdali), hervor. Dieses Toxin aktiviert die Kalium-Pumpe. Bei Fusicoccineinwirkung auf Spaltöffnungen bleiben diese offen, der Wasserverlust kann höher als der Nachschub sein, das Wasserdefizit führt zum Welken. Der biologische Vorteil für den Pilz liegt im Vorfinden offener Spaltöffnungen, denn das sind, außer Wundstellen, die einzigen Orte, durch die seine Hyphen in das Blattgewebe eindringen können. Sofern die Fusicoccinwirkung dabei lokal eingeschränkt bleibt (es genügt im Prinzip eine Spaltöffnung für eine Pilzhyphe), wird der Gesamtwasserhaushalt zunächst kaum oder gar nicht beeinflußt.
Bei Wasserdefizit produziert die Pflanze in verstärktem Maße Abscisinsäure, die aktiv zu den Schließzellen transportiert und in ihnen gespeichert wird. Sie wiederum hemmt die Kalium-Ionenpumpe, verhindert damit den Aufbau eines osmotischen Drucks und bewirkt Spaltenverschluß.
Zusammenfassend können wir feststellen, daß das Öffnen und Schließen über zwei voneinander unabhängige Regelkreise (den Wasser- und den Kohlendioxyd-Regelkreis) gesteuert wird. Die Regulation über das Wasserpotential ist der wirkungsvollere Mechanismus. Dabei wird einmal die Wassermenge in unmittelbarer Nachbarschaft der Schließzellen verrechnet, und zum anderen, über die ABA-Wirkung, das Wasserpotential in entfernt gelegenen Gewebeabschnitten. Bei geschlossenen Spaltöffnungen können Wasser- und Kohlendioxyd-Regelkreis miteinander konkurrieren, denn in photosyntheseaktiven Geweben ist Kohlendioxyd in der Regel ein limitierender Faktor. Bei gleichzeitigem Wassermangel bleiben die Spaltöffnungen dennoch geschlossen, die Photosyntheserate sinkt auf ein niedriges Niveau, kommt aber nicht ganz zum Erliegen, da Kohlendioxyd durch Atmungsprozesse innerhalb der Pflanze in nicht unbeträchtlicher Menge immer wieder neu gebildet wird.
|
|