
Es gibt eine Anzahl von Pflanzenarten, die sich bei hoher Lichtintensität gegenüber den übrigen durch eine erhöhte und weit effizientere Nettophotosyntheseleistung auszeichnen als die übrigen. Paradebeispiele hierfür sind etliche Gramineenarten wärmerer Gegenden, wie z.B. Mais und Zuckerrohr.
Anfang der sechziger Jahre stellte H. KORTSCHAK (Hawaiian Sugar Planter's Association) fest, daß das erste Produkt der Photosynthese beim Zuckerrohr nicht die C3-Verbindung 3-Phosphoglycerat, sondern eine Verbindung aus vier C-Atomen ist. Die Pflanzenphysiologen, der Australier M. D. HATCH und der Engländer C. R. SLACK bestätigten den Befund und identifizierten die Verbindung als Oxalacetat (oxal acetic acid: OAA), das durch Anlagerung eines Kohlendioxyd-Moleküls an Phosphoenolpyruvat (PEP) entsteht (HATCH-SLACK-Zyklus = C4-Zyklus). Pflanzenarten, bei denen dieser Weg beschritten wird, nennt man C4-Pflanzen (respektive CAM-Pflanzen), im Gegensatz zu den C3-Pflanzen, bei denen das aufgenommene Kohlendioxyd direkt in den CALVIN-Zyklus eingebracht wird. Das Oxalacetat wird in der Mehrzahl der Fälle in Malat überführt, aus dem Kohlendioxyd enzymatisch wieder abgespalten wird.
Dieses Kohlendioxyd wird nunmehr dem Ribulose-1,5-Diphosphat zugeführt und via CALVIN-Zyklus fixiert.
Anstelle von Malat tritt bei einigen Arten Aspartat als Zwischenprodukt auf:
Oxalacetat + L-Glutamat > Aspartat + alpha-Ketoglutarat.
Die reversible Bindung des Kohlendioxyds dient ganz offensichtlich seiner Akkumulation und Speicherung. Da der Prozeß energieverbrauchend ist, kann man von einer Kohlendioxyd-Pumpe sprechen.
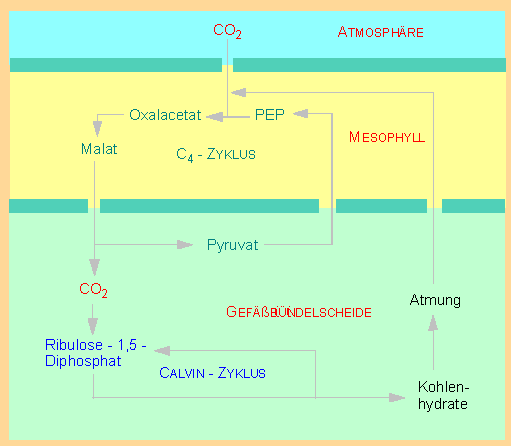
Photosynthese in C4-Pflanzen. In Mesophyllzellen wird CO2 an Phosphoenolpyruvat (PEP) gebunden. Es entsteht Oxalacetat und daraus Malat. Dieses wird an die Zellen der Gefäßbündelscheide ("Kranz"-Zellen) abgegeben, CO2 wird abgespalten und dem CALVIN- Zyklus zugeführt. Das Pyruvat wandert in die Mesophyllzellen zurück (aktiver Transport unter Energieverbrauch) und wird unter zusätzlichem ATP-Verbrauch zu PEP phosphoryliert
Die Blätter der C4-Pflanzen (mit sog. "Kranz"- Zellen) unterscheiden sich anatomisch grundsätzlich von denen der C3-Pflanzen. In C3-Pflanzen sind die Chloroplasten einheitlich strukturiert, in C4-Pflanzen kommen zwei Chloroplastentypen vor. Die Mesophyllzellen enthalten normal ausgebildete Chloroplasten, während die Zellen der Gefäßbündelscheide granalose und damit nur partiell funktionsfähige Chloroplasten enthalten. Die Beeinträchtigung bezieht sich jedoch nur auf die Lichtreaktionen der Photosynthese. Der Calvin-Zyklus ist hiervon nicht betroffen. Die primäre Kohlendioxyd-Bindung (HATCH-SLACK-Reaktion) erfolgt in den Mesophyllzellen, der Einbau in Kohlenhydrate (CALVIN-Zyklus) in den Zellen der Gefäßbündelscheide. Die beiden Teilabschnitte der Photosynthese sind damit räumlich voneinander getrennt.
CAM ist die Abkürzung für Crassulacean-Acid-Metabolism. Die Benennung weist darauf hin, daß dieser Stoffwechselweg vornehmlich bei den Crassulaceae (und anderen Sukkulenten) auftritt. Die chemische Reaktion der Kohlendioxyd-Anreicherung gleicht der in C4-Pflanzen, doch sind hier die beiden Teilabläufe nicht räumlich, sondern zeitlich voneinander getrennt. CAM-Pflanzen kommen vornehmlich in Trockengebieten vor. Ein Öffnen der Stomata zur Kohlendioxyd-Gewinnung ist dort stets mit einem besonders hohen Wasserverlust verbunden. Um ihn bei starker Sonneneinstrahlung einzudämmen bzw. fast ganz zu unterbinden (die kutikuläre Transpiration bleibt erhalten), hat sich ein Regelmechanismus entwickelt, der eine nächtliche Kohlendioxyd-Aufnahme ermöglicht. Das vorfixierte Kohlendioxyd wird als Malat (und Isocitrat) in Vakuolen gespeichert und tagsüber für die Photosynthese genutzt.
Das Enzym, das die primäre Kohlendioxyd-Fixierung der C4-
und CAM-Pflanzen katalysiert, ist die Phosphoenol- Pyruvatcarboxylase (PEPC),
deren Affinität zum Kohlendioxyd weit höher ist als die entsprechende
Affinität der RuDP-Carboxylase (Rubisco), dem ersten Enzym des CALVIN-Zyklus.
C4-Pflanzen werden somit in die Lage versetzt, auch noch Spuren
von Kohlendioxyd zu nutzen. PEPC kommt in geringer Menge (ca. 2 - 3%) auch
in C3-Pflanzen vor, und auch dort fällt ihr eine Schlüsselrolle
bei der Regulation des Stoffwechsels zu.

Kohlenstoffgewinn bei C4 - und C3 -Pflanzen offener Graslandschaften in verschiedenen geographischen Breiten. In der gemäßigten Zone ist die geringe Lichtintensität für eine Benachteiligung von C4 -Pflanzen ausschlaggebend, C3 -Pflanzen sind im Vorteil, weil die Photorespirationsrate niedrig ist und keine Energie für die Vorfixierung von CO2 eingesetzt zu werden braucht. (J. R. EHRLICHER, 1978).
Unter anderem sorgt sie in wachsenden Wurzeln (z.B. von Maiskeimlingen) für die Bereitstellung von NADH + H+, das zur Lipidsynthese benötigt wird. Dabei laufen die folgenden Reaktionen ab:
- Phosphoenolpyruvat + HCO3- > Oxalacetat + Pi
- Oxalacetat > Malat. - Während dieses Schritts wird NADH + H+ zu NAD+ oxydiert.
- Malat > Pyruvat + CO2.
Hierbei erfolgt die Reduktion von NAD zu NADH + H+.
In den Wurzelknöllchen der Leguminosen erfolgt eine Stickstoff-Fixierung. Zum Einbau des durch bakterielle Aktivität erzeugten Ammoniaks müssen Kohlenstoffskelette in ausreichender Menge bereitgestellt werden. Die CO2-Fixierung via PEPC-reaction dient daher auch als wichtige Ergänzung zum Auffüllen des Kohlenstoffskelettvorrats
Ferner dient die PEPC zur Produktion von Intermediärprodukten des Citratzyklus (Oxalacetat und/oder Malat), um ggf. Engpässe zu überwinden. PEPC-Aktivitäten unterliegen der Kontrolle durch externe Faktoren, wobei die Tageslänge eine der ausschlaggebenden Größen ist. In einigen untersuchten Fällen sind in unterschiedlichen Geweben verschiedene Isoenzyme nachgewiesen worden, deren Produktion durch unterschiedliche Auslöser kontrolliert wird.
In C3-Pflanzen kann bei hoher Strahlungsintensität bis zu 20 Prozent des im CALVIN-Zyklus fixierten Kohlenstoffs durch Photorespiration wieder verlorengehen. Bei hoher Lichtintensität ist die Photorespiration etwa 1,5 - 3,5mal so hoch, wie die normale Dunkelatmung. In C4-Pflanzen ist die Photorespiration drastisch reduziert, vielleicht ist sie überhaupt nicht mehr nachweisbar. Mit anderen Worten:
Die Nettophotosyntheserate (und damit die Nettoproduktion an Biomasse) der C4-Pflanzen liegt bei hohen Lichtintensitäten weit höher als die der C3-Pflanzen. Das Temperaturoptimum der Photosynthese liegt unter dem der Dunkelatmung. Folglich spielen Verluste durch Atmung bei höheren Temperaturen eine größere Rolle als bei niedrigen. Wo Lichtmangel ein limitierender Faktor ist und die Temperaturen niedrig liegen (in gemäßigten Klimazonen), sind C3-Pflanzen im Vorteil, C4-Pflanzen treten kaum auf (eine der Ausnahmen ist Spartina townsendii). C4-Pflanzen (es sind fast immer Kräuter oder Sträucher) sind im offenen Gelände wärmerer Klimazonen erfolgreicher.
Es sei vermerkt, daß im HATCH-SLACK-Zyklus pro fixiertem Kohlendioxyd-Molekül zwei ATP Moleküle verbraucht werden.
C4-Pflanzen gehören zahlreichen, phylogenetisch nicht zusammenhängenden Familien der Mono- und der Dikotyledonen an. Darüber hinaus fand man C4-Aktivitäten u.a. auch bei der Blaualge Anacystis nidulans sowie bei einigen Dinoflagellaten.
Da bei den höheren Pflanzen mit der Alternative C3 oder C4 beträchtliche anatomische Veränderungen der Blätter verbunden sind, muß man davon ausgehen, daß das genetische Potential zur Realisierung beider Wege im Pflanzenreich verbreitet ist und daß in Abhängigkeit von ökologischen Ansprüchen bei einer Art der eine, bei einer verwandten der andere Weg eingeschlagen wird.
Ein vielstudiertes Beispiel stellt die Gattung Atriplex dar, bei der beide Wege verwirklicht sind. Demnach gehören C3-Arten einer Verwandtschaftsgruppe, C4-Arten einer anderen an. In Einzelfällen lassen sich Bastarde zwischen C3- und C4-Arten herstellen.
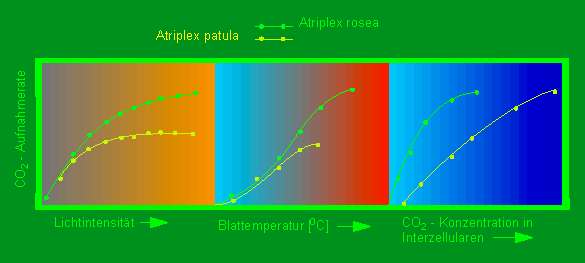
Einfluß verschiedener Parameter auf die Effizienz der Kohlendioxyd-Aufnahme (Ordinate) bei einer C3 - Pflanze (Atriplex patula - gelbe Linien) und einer C4 - Pflanze (Atriplex rosea - grüne Linien). Gemessene Parameter von links nach rechts: Lichtintensität, Blattemperatur und Kohlendioxyd-Konzentrationen in den Interzellularen (Nach O. BJÖRKMAN und J. BERRY, 1973)
Bei mehreren Pflanzenarten aus den Gattungen Zea, Mollugo, Moricandia, Flaveria u.a. werden in einer Pflanze beide Wege beschritten. In jungen Blättern erfolgt die Photosynthese meist auf dem C3-, in älteren auf dem C4-Weg. Der Betrag des C4-Anteils wird durch Standortfaktoren reguliert.
CAM: Vor- und Nachteile. CAM wurde bei über 1000 Angiospermen aus 17 verschiedenen Familien nachgewiesen. In der Regel ist er mit Sukkulenz der Pflanzen verknüpft, aber nicht alle Crassulaceae z.B. zeigen CAM, und Sukkulenz ist nicht die Vorbedingung für CAM. Die Bromeliacee Tillandsia usneoides ist nicht sukkulent, zeichnet sich aber durch CAM aus. Mesembryanthemum crystallinum (eine Blattsukkulente) kann den C3-Weg einschlagen, schaltet aber bei Wachstum auf salzhaltigen Böden auf CAM um. Experimentell kann die Umschaltung durch Erhöhung des NaCl-Gehalts des Nährmediums erreicht werden (K. WINTER und D. J. von WILLERT, 1972). Während der Vorteil der C4 Pflanzen bei hohen Lichtintensitäten zum Tragen kommt, regulieren bei CAM-Pflanzen vornehmlich Temperatur, Luftfeuchtigkeit und Salinität den Grad des CAM-Einflusses. Man kennt starke und schwache CAM-Pflanzen. Bei den schwachen tritt CAM nur bei bestimmten Differenzen von Tag- und Nachttemperaturen in Erscheinung. CAM-Pflanzen, die viel Malat und wegen des dadurch bedingten osmotischen Wertes auch viel Wasser gespeichert haben, sind in der Regel weniger frostresistent als C3-Pflanzen. Sie sind wegen des hohen Säuregehalts aber auch weniger hitzeresistent. Für Arten in ariden (trockenen) Gegenden ist es daher notwendig, tagsüber den Malatpool abzubauen (R. LÖSCH und H. KAPPEN, Universität Kiel, 1985). Im allgemeinen schließen der C4-Weg und CAM einander aus. Als Ausnahme wird die sukkulente C4-Dikotyledone Portulaca oleracea genannt, die an natürlichen Standorten den jeweils optimalen Weg einschlagen kann.
|
|