"Es betrifft die Tatsache, daß gewisse Baumarten ganz regelmäßig sich im Boden nicht selbständig ernähren, sondern überall in ihrem gesamten Wurzelsystem mit einem Pilzmycelium in Symbiose stehen, welches ihnen Ammendienste leistet und die ganze Ernährung des Baumes aus dem Boden übernimmt. . . Dieser Pilzmantel hüllt die Wurzel vollständig ein auch den Vegetationspunkt derselben lückenlos überziehend, er wächst mit der Wurzel an der Spitze weiter und verhält sich in jeder Beziehung wie ein zur Wurzel gehörendes, mit dieser organisch verbundenes peripheres Gewebe. Der ganze Körper ist also weder Baumwurzel noch Pilz allein, sondern ähnlich wie ein Thallus der Flechten eine Vereinigung zweier verschiedener Wesen zu einem einheitlichen morphologischen Organ, welches vielleicht passend als Pilzwurzel, Mykorrhiza bezeichnet werden kann... Dieser (der Mantel) liegt der Wurzelspitze nicht bloß innig auf, sondern von ihm aus dringen Pilzfädchen auch zwischen den Epidermiszellen in die Wurzel selbst ein... (doch) nie wurden die Fäden bis zur Endodermis verfolgt... Sie treten nie in das Lumen der Zellen ein." (B. FRANK, 1885)
115 Jahre Mykorrhizaforschung förderten zutage, daß nahezu 80 Prozent aller Landpflanzen Mykorrhiza-Assoziationen ausbilden können. Man unterscheidet dabei zwischen zwei grundsätzlich voneinander verschiedenen Typen:
- ektotrophe Mykorrhiza (kurz Ektomykorrhiza)
- endotrophe Mykorrhiza (oder Endomykorrhiza)
Zu den Endomykorrhizen gehört auch die sogenannte Vesikular-arbuskuläre Mykorrhiza (kurz VA-Mykorrhiza), die sich inzwischen als der am weitesten verbreitete Typ herausgestellt hat .
Unter Ektomykorrhiza versteht man genau das, was durch das FRANKsche Zitat wiedergegeben wurde. Der Pilz formt eine Hülle (einen Mantel) um die Wurzelspitzen. Die Pilzhyphen dringen zwar in das Rindengewebe der Wurzel ein, nicht jedoch in die Zellen.
Bei der Endomykorrhiza werden Haustorien ausgebildet. Ein Teil des Pilzmycels breitet sich in und zwischen den Rindenzellen aus, ein anderer erstreckt sich in das Erdreich hinein. Ein zusammenhängender Mycelmantel wird in der Regel nicht gebildet. Die VA-Mykorrhiza zeichnet sich durch spezifisch geformte Haustorien aus. Im Zellumen bilden sich Vesikel oder büschelförmig verzweigte Gebilde (Arbuskeln). Die Hyphen der VA-Mykorrhizapilze sind septenlos, d.h., sie besitzen keine Zellquerwände. Sie gehören ausnahmslos (?) zur Pilzklasse der Zygomyceten. Die Pilze der Ektomykorrhiza und der Nicht-VA-Endomykorrhiza sind vorwiegend Basidiomyceten, seltener Askomyceten (die Hyphen dieser Pilze enthalten Septen); als Ektomykorrhizapilze sind gelegentlich auch Zygomyceten nachgewiesen worden.
Ausführliche Darstellung aller Mykorrhizatypen:
Ektomykorrhiza findet man bei einer Reihe von Baum- und Straucharten, vornehmlich aus den Familien Pinaceae, Cupressaceae, Fagaceae, Betulaceae, Salicaceae, Dipterocarpaceae, Myrtaceae und Caesalpinaceae. Die Mehrzahl der Mykorrhizabäume kommt in den borealen (kalten) und gemäßigten Klimazonen oder an nährstoffarmen Standorten der Tropen vor. Wie schon FRANK richtig erkannte, dient der Pilz dem Baum, indem er ihn mit in Wasser gelösten Mineralien versorgt. Darüber hinaus zeigte sich, daß der Pilzmantel auch einen Schutz vor parasitischen Erdpilzen (Hallimasch, Wurzelfäule, Phytophthora u.a.) bietet. Der Schutz beruht in einigen Fällen auf einer von der Pflanze erworbenen, durch den Pilz induzierten Resistenz, die an der Produktion von Abwehrstoffen (phenolischen Verbindungen) erkennbar ist. Mykorrhizapilze sondern oftmals Phytohormone aus, die das Wurzelwachstum beeinflussen. Das Wurzelsystem und Pilzmycel sind zusammen für die Ausbildung einer Rhizosphäre verantwortlich, in der sich die Lebensbedingungen von denen in der Umgebung unterscheiden; so ist die Rhizosphäre fast immer bakterienreicher als die übrigen Bodenregionen.
Den Mykorrhizapilzen stellt die Pflanze Kohlenhydrate (meist Glucose), Wachstumsstimulatoren (flüchtige Terpene) und Vitemine (Thiamin oder dessen Vorstufen, Pyrimidin oder Thiazol, Biotin, Pantothenat, Nicotinsäure u.a.) zur Verfügung. Viele der Pilze sind vitaminheterotroph, so daß sie auf eine Versorgung durch die Pflanzen angewiesen sind. Das Bedarfspektrum ist jedoch von Art zu Art unterschiedlich. Viele Mykorrhizapilze können isoliert und in Kultur genommen werden, doch nur selten werden unter diesen Bedingungen Fruchtkörper gebildet.
Die eine Fruchtkörperbildung stimulierenden Faktoren sind in den meisten Fällen bislang nicht bestimmt worden. In der Regel sind Mykorrhizapilze gegenüber Überwachsung durch andere (saprophytische) Pilze extrem empfindlich. Zu den Mykorrhizabildnern gehören bekannte Basidiomyceten (wie Steinpilz, Knollenblätterpilz, Täubling, Kartoffelbovist u.a.) sowie Askomyceten (z.B. die Trüffeln). Weltweit sind etwa 5000 ektomykorrhizabildende Arten identifiziert worden. In bezug auf den Baumwirt sind sie im allgemeinen wenig wählerisch. Junge Bäume sind öfter mit anderen Pilzen assoziiert als ältere. Etliche Pilzarten, wie beispielsweise der Fliegenpilz, wurden in Assoziation mit einer Anzahl von Baumarten gefunden; nur wenige, z.B. den Lärchenröhrling, findet man ausschließlich in Lärchenwäldern.
Ein zusammenhängendes Mycel verbindet vielfach die Wurzelsysteme benachbarter Bäume der gleichen oder verschiedener Arten (s.a. folgenden Abschnitt). Die Ektomykorrhiza ist für eine Samenkeimung und für die frühen Entwicklungsstadien der Baumkeimlinge entbehrlich. Unter natürlichen Bedingungen wurde bei nahezu allen wenige Monate alten Keimlingen Pilzassoziationen gefunden. Unter kontrollierten Bedingungen ließ sich im Experiment belegen, daß Keimlinge in steriler Erde wesentlich schlechter gediehen als jene mit Mykorrhizapilzen .
Morphologisch unterscheiden sich die infizierten Wurzeln von nicht infizierten. Die Rindenzellen bilden keine Sekundärwände aus, Wurzelhaare und Wurzelhauben fehlen, die Endodermis ist meist zusätzlich durch Tannine imprägniert, die ein weiteres Vordringen der Pilze verhindern. Das Hyphennetz im Rindengewebe wird HARTIGsches Netz genannt.
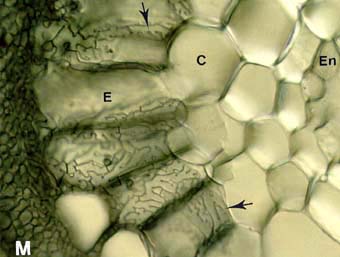
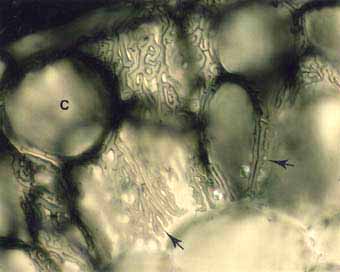
Links: Pinus strobus: Ektomykorrhiza in der Wurzel. HARTIG Netz Hyphen (Pfeile) entwickeln sich an den Oberflächen der Rindengewebszellen (Cortex =C). - Rechts: Populus tremuloides: Ektomykorrhiza - Wurzelquerschnitt mit labyrinthähnlichen HARTIG Netzhyphen (Pfeile), die die länglichen Epidermalzellen umgeben. Der Verzweigungsgrad des Pilzmycels erhöht damit den Kontaktbereich zu den Pflanzenzellen.
Im Gegensatz zu den saprophytisch oder parasitisch lebenden Pilzen sezernieren die meisten Mykorrhizapilze keine Phenoloxydasen (für Ligninabbau erforderlich) und keine Cellulasen (für Celluloseabbau erforderlich). Das gilt allerdings nur, solange ihnen ausreichende Glucosemengen angeboten werden. Sobald die Zufuhr stockt, wird Cellulase synthetisiert, und der bis dahin symbiotisch lebende Pilz geht zu parasitischer Lebensweise über. Andererseits ist für zwei Mykorrhizapilzarten erwiesen, daß sie ständig große Mengen an Cellulase, Xylanase, Amylase und Proteinase (jedoch keine Pektinase) produzieren, ohne daß die Symbiose darunter leidet. Mykorrhizapilze bevorzugen leicht saure Böden; pH-Werte zwischen 4,0 und 5,0 sind optimal. Oberhalb von pH 7 können sie nicht existieren. In Calcium-reichen Böden (z.B. in Kalkbuchenwäldern) sind Mykorrhizen extrem selten, bzw. nur rudimentär ausgebildet. Dieser Befund ist im Zusammenhang mit den Hilfsmaßnahmen gegen die Schäden von Saurem Regen nennenswert, denn durch die Kalkung von Waldböden wird der pH-Wert lokal drastisch erhöht, was wiederum zu erheblichen Schäden der Mykorrhizapilze und sekundär zu weiteren Schäden der Waldbäume führen kann.
Zu den beiderseitigen Vorteilen der Mykorrhiza gehören die bereits genannte effizientere Nutzung des Nährstoffangebots. Da sich das Pilzmycel gegenüber dem Wurzelsystem durch eine um ein Vielfaches größere Oberfläche auszeichnet, kann ein größeres Bodenvolumen erschlossen werden. Zum Aufbau eines ebenso komplexen Wurzelsystems (ohne Pilzmycel) müßte die Pflanze weit mehr Energie investieren, als ihr durch Abgabe von Kohlenhydraten an den Pilz verlorengeht. Kohlenhydrate wiederum sind für diesen essentiell, denn Pilze decken ihren Stickstoffbedarf vornehmlich durch Aufnahme reduzierter Stickstoffverbindungen (Ammonium-, Aminogruppen), für deren Fixierung Akzeptormoleküle benötigt werden (Kohlenstoffskelette). Ein Mangel an Kohlenhydraten hemmt die Ausprägung einer Mykorrhiza, ein Mangel an Stickstoff oder Phosphor fördert sie.
Pilze sezernieren Protonen und säuern damit den Boden an. Im Gegenzug (durch Aufbau eines elektrochemischen Potentials) nehmen sie größere Mengen an Phosphat auf und akkumulieren es intrazellulär als Polyphosphat. Ein beträchtlicher Teil davon kommt der Pflanze zugute. Wie Meßwerte veranschaulichen, sind bei Mykorrhizakeimlingen - im Vergleich zu den Kontrollen - vor allem die Phosphatwerte, weniger die Stickstoff- und Kaliumwerte erhöht.
Wie wichtig die Anwesenheit der "richtigen" Mykorrhizapilze für ein Baumwachstum ist, ergibt sich u.a. aus der Erfahrung, daß Grasland (Prärie), das weitgehend frei von diesen Arten ist, nur schwer aufzuforsten ist. Zahlreiche Versuche sind gescheitert. Erst nach Inokulation der Bäume (in Baumschulen) gelangen die Aufforstungsmaßnahmen.
Die Abgrenzung von Ekto- und Endomykorrhiza ist keineswegs so scharf und eindeutig, wie man es vielleicht nach den eingangs präsentierten Definitionen annehmen würde. Es gibt zahlreiche fließende Übergänge, und vielfach geht eine Ektomykorrhiza in eine Endomykorrhiza über. Für solche Fälle wurde der Begriff Ekto-Endomykorrhiza geprägt. Wie schon erwähnt, ist die VA-Mykorrhiza der bei weitem häufigste Typ. Bevor Einzelheiten über sie beschrieben werden, müssen zwei andere Erscheinungsformen, nämlich die Mykorrhiza der Ericales und die der Orchideen erwähnt werden.
Ericales, vor allem Arten aus der Familie der Ericaceen, sind in der Natur stets mit Pilzen assoziiert, sie gelten daher als obligat mycotroph. Das heißt aber nicht, daß man Ericaceen nicht pilzlos kultivieren könnte. Unter Kulturbedingungen, auf rein anorganischem Substrat, wachsen Calluna, Vaccinum, Azaleen u.a. ebenso gut wie in der Natur zusammen mit Pilzen. Setzt man dem Nährmedium jedoch organisches Material (z.B. Pepton oder Hefeextrakt) zu, ist das Wachstum stark gehemmt. Offensichtlich sezernieren die Wurzeln Substanzen, die in Reaktion mit dem organischen Material Toxine produzieren. In Anwesenheit von Pilzen werden jene wieder inaktiviert. Mit anderen Worten: das Wachstum ist auf nährstoffreichen Böden mit Mykorrhizapilzen optimal. Aber gerade die Ericaceen sind in der Natur fast nur auf sauren, extrem nährstoffarmen Böden zu finden. Der Vorteil der obligaten Assoziation mit Pilzen erlaubt es ihnen, auch diese Böden effizient auszubeuten. Der Pilzbefall erfolgt bei einer Reihe von Arten kurz oberhalb des Vegetationspunkts. Bei Calluna jedoch wird durch den Pilz der primäre Vegetationspunkt der Wurzel zerstört, und als Folge davon werden sekundäre Vegetationszonen aktiviert, was wiederum zu einer Steigerung der Anzahl an Verzweigungen im Wurzelsystem führt, und damit zu einer besseren Durchdringung des Bodens.
In der Forstwirtschaft hat es sich als problematisch erwiesen, Calluna-Flächen aufzuforsten. Ein Grund dafür ist das Fehlen jener Mykorrhizapilze, auf die Waldbäume angewiesen sind. Das Fehlen beruht auf einer Produktion von Hemmstoffen der mit Calluna vergesellschafteten Mykorrhizapilze. Experimentel ließ sich zeigen, daß Extrakte aus Calluna-Humus das Mycelwachstum vieler Pilze hemmen. Nur einige wenige Arten, wie Boletus scaber und Amanita muscaria, sind hiergegen resistent. Beide Pilzarten werden überall dort als Mykorrhizapilze gefunden, wo Birken in Calluna-Flächen eindringen.
Eine der Ericaceen-Mykorrhiza ähnliche Erscheinung findet man beim Fichtenspargel (Monotropa), einer chlorophyllfreien, saprophytisch lebenden Pflanze sowie bei verschiedenen Arten der (grünen) Pyrolaceen. Monotropa ist ein Wurzelparasit, der auf verschiedenen Laub- und Nadelbäumen gedeihen kann. Seine Wurzel ist von einem dichten Pilzmycel umhüllt, dessen Ausläufer sowohl das umgebende Erdreich als auch die Wurzeln der Wirtsbäume durchdringen und damit einen indirekten Kontakt zwischen Wirt und Parasit herstellen. Daß durch solche Brücken tatsächlich Assimilate fließen, belegten R. FRANCIS und D. J. READ (1984) experimentell am Beispiel einer VA-Mykorrhiza.
Sie pflanzten radioaktiv markierte Keimlinge von Plantago major zusammen mit nichtmarkierten von Festuca ovina in ein Kulturgefäß, in einem weiteren Experiment wurde Plantago mit einem Pilz inokuliert. Die Wurzelsysteme beider Arten sind morphologisch leicht voneinander unterscheidbar. Im pilzhaltigen Kulturgefäß breitete sich die Radioaktivität nach kurzer Kulturdauer sowohl im Pilz als auch in Festuca ovina aus. In der pilzfreien Kultur blieben die Festuca ovina-Wurzeln trotz direktem physischem Kontakt mit den markierten Plantago-Wurzeln unmarkiert .
Wie die Ericaceen sind auch die Orchidaceen obligat mycotroph, doch liegen die Ursachen dafür hier woanders (H. BURGEFF, 1909, 1936). Orchideensamen sind extrem klein (0,3-14 µm), in der Regel sind keine Kotyledonen vorhanden, ein Same kann zwar keimen, sich aber nicht über ein Wenigzellstadium hinweg entwickeln. Nur in Assoziation mit Pilzen, die hier das Nährsubstrat stellen, ist eine Weiterentwicklung möglich. Orchideen sind daher, zumindest in ihrer ersten Lebensphase, Parasiten. Viele von ihnen (jene mit grünen Blättern) gehen in einem späteren Entwicklungsstadium zu autotropher Ernährung über. Von diesem Zeitpunkt an ist der Pilz überflüssig.
Der Mykorrhizapilz dringt (meist durch den Suspensor) in das Gewebe des jungen Keimes ein und breitet sich von dort in die entstehenden Wurzeln aus. Sproß und Wurzelknollen (soweit vorhanden) sind in der Regel pilzfrei.
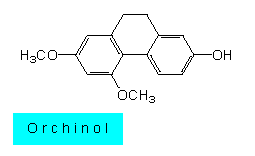
Die endotroph lebenden Pilze gehen im Verlauf der pflanzlichen Entwicklung meist zugrunde, die Pilzreste werden von den Orchideen resorbiert. Unterbleibt diese wirtsspezifische Aktion (oft bereits in recht frühen Entwicklungsstadien), breitet sich der Pilz aus und wird parasitisch. Bei zahlreichen Orchideenarten kann sich daher nur ein geringer Prozentsatz der Keimlinge fortentwickeln. Die Hemmung des Pilzwachstums beruht auf der Synthese eines Antagonisten, der zunächst als Orchinol bezeichnet wurde und den E. GÄUMANN und H. KERN (1959) als Dehydroxyphenanthrin charakterisierten. Dieses Fungizid wirkt auf zahlreiche Mykorrhiza- und Erdpilze. Seine Synthese wird nur bei Pilzanwesenheit induziert.


Reife Arbuskeln von Glomus mosseae mit zahlreichen fein verzweigten Hyphen. - Arbuskel von Gigaspora margarita mit verlängerter Stammhyphe (trunk hypha = T) und Aggregationen feiner verzweigter Hyphen (Pfeile). Man beachte die Unterschiede zu den Glomus Arbuskeln (links).
Lappige Vesikel einer Acaulospora Art in der Wurzel einer Kleeart.
Die VA-Mykorrhiza ist die verbreitetste und in den letzten Jahren am intensivsten studierte Mykorrhizaform. Es gibt keine Pflanzenfamilie, bei der sie nicht gefunden wurde; lediglich bei den Juncaceae, Cyperaceae, Caryophyllaceae und Brassicaceae ist sie selten. Bei Pflanzen feuchter Standorte ist sie weniger stark ausgeprägt als bei jenen trockener. Allerdings sind Pflanzen der Salzwiesen oft stark verpilzt. Die Pilze durchsetzen das Rindengewebe der Wurzel, doch nie Meristeme oder Leitgewebe sowie chlorophyllhaltige oder andersartig spezialisierte Gewebe.
Ein wesentliches Merkmal der bereits besprochenen Ektomykorrhiza ist die hohe Artenzahl der Pilze. Fiir die VA-Mykorrhiza gilt das Gegenteil, weltweit sind nur etwa 30 morphologisch voneinander unterscheidbare Formen (Arten?) identifiziert worden. Die Mehrzahl von ihnen wird zur Gattung Glomus gerechnet. D. W. MALLOCH, K. A. PIROZYNSKI und P. H. RAVEN wiesen 1980 darauf hin, daß die VA-Mykorrhiza vornehmlich in artenreichen Pflanzengesellschaften vorkommt, Ektomykorrhiza hingegen in artenarmen. Die Zahl der Pflanzenarten verhält sich damit umgekehrt proportional zur Zahl der symbiotischen Pilze.
Die Ektomykorrhiza trat vermutlich erst in der Mittleren Kreide auf, sie ist als eine progressivere Form der Symbiose einzustufen, die es den Baumarten ermöglichte, in gemäßigten und kalten Klimazonen oder auf armen Böden zu bestehen. Die VA-Mykorrhiza ist die primitivere Form der Symbiose. Es gibt sie, seitdem es vielzellige Landpflanzen gibt. Durch Fossilfunde aus dem Devon wurden Assoziationen zwischen Pilz und Rhynia sowie Asteroxylon nachgewiesen. Auch die heute existierenden primitivsten Gefäßpflanzen (Psilotum und Tmesipteris) zeichnen sich durch VA-Mykorrhiza aus. Da jene bei Holzpflanzen weiterverbreitet als bei Kräutern ist, lag der Gedanke nahe, daß sich große Bäume ohne sie überhaupt nicht haben entwickeln können. Wie schon dargelegt, gilt auch hier, daß das Mycelsystem die resorbierende Oberfläche im Boden um ein Vielfaches erhöht. Durch Isotopenversuche konnte gezeigt werden, daß 32P sich im Boden in einem gegebenen Zeitraum durch Diffusion um ca. 7 mm ausbreitet. In pilzdurchsetzten Böden beträgt die Ausbreitung im gleichen Zeitraum 7,5 cm.
Einer der größten Vorteile auch der VA-Mykorrhiza liegt in der Zusatzversorgung der Pflanzen mit Phosphat. Die Aussage ist durch zahlreiche Experimente abgesichert. Dabei konnte aber auch gezeigt werden, daß extrem hohe Phosphatgehalte [1 g Calciumphosphat pro kg Erde] die Ausbildung des Mycelsystems hemmen.
Pflanzenarten (Kräuter) mit gut ausgeprägtem Wurzelhaarsystem gehen nur an phosphatarmen Standorten eine Symbiose mit Pilzen ein. Bei Wassermangel ist die Photosyntheserate (Kohlendioxyd-Fixierung) von Pflanze-Pilz-Assoziationen deutlich höher als die von pilzfreien Exemplaren der gleichen Pflanzenart. In bezug auf die von der Pflanze bereitgestellten Kohlenhydrate sind die VA-Mycorrhizapilze wenig wählerisch. Sie sind befähigt, ein weites Spektrum an Mono-, Di- oder Polysacchariden zu verwerten.
|
|