
Von besonderem Interesse ist dabei die Frage, ob Proteine und Nukleinsäuren entstehen können.
S. W. FOX (University of Miami) hat sich seit den sechziger Jahren mit der Bildung von Proteinen aus Aminosäuren befaßt. Seine Untersuchungen ergaben, daß Proteine und/oder proteinähnliche Polymere (Proteinoide) bei einer Erhitzung eines Aminosäuregemisches auf 150-200 Grad C entstehen. Die Inkubationszeit beträgt mehrere Tage. Unter Zusatz von Polyphosphat steigt die Ausbeute, die Polymerisation erfolgt bereits bei 70 Grad C. Damit stellt sich für uns sofort die Frage nach den Leistungen dieser Polymere, und darauf gibt es zwei, für alles Weitere entscheidende Antworten:
Proteinoide entfalten katalytische Aktivitäten. Es werden zwar noch nicht alle Eigenschaften moderner Enzyme erreicht, vor allem nicht deren Reaktionsgeschwindigkeit, es wird aber deutlich, daß die Voraussetzungen für eine Vervollkommnung durch eine Evolution gegeben sind. Aus der Kinetik einer Reaktion ist ablesbar, daß Proteinoide bereits kooperatives Verhalten (allosterisches Verhalten) zeigen.
Proteinoide sind strukturbildend. Sie haben die Tendenz zur Aggregation (Quartärstrukturbildung). In wäßriger Lösung entstehen hohlkugelförmige Gebilde (Mikrosphären), deren Bildung wiederum ein wichtiger Schritt in Richtung Zellentstehung ist. Vorsichtig gesagt, könnte man Mikrosphären als Vorstufen von Protozellen (Vorstufen der Organisationsform Zelle) ansehen. Eine wichtige Eigenschaft haben sie mit den Zellen gemeinsam: Sie bilden eine Hülle, durch die ein Reaktionsvolumen von der Umwelt abgetrennt wird.
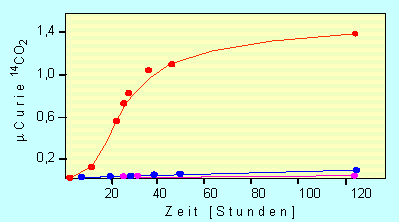
Die katalytische Aktivität eines Proteinoids. Gemessen wurde die Dekarboxylierungsrate von Brenztraubensäure (Pyruvat) in Anwesenheit eines Proteinoids (rot), eines Gemisches aus freien Aminosäuren (blau) und ohne irgendwelchen Zusatz (magenta)
(Nach S. W. FOX und K. DOSE, 1972)
Self-assembly (Selbstorganisation) ist demnach eine inhärente Eigenschaft der Materie. Sie beruht u.a. auf der Tendenz hydrophober Molekülanteile, unter Wasserausschluß die maximal mögliche Zahl schwacher Wechselwirkungen auszubilden. Bei der Analyse der Proteinoidzusammensetzung wurde deutlich, daß die Polymerisation nicht allein den Zufallsgesetzen unterworfen ist. Die Aminosäurezusammensetzung unterscheidet sich nämlich ganz erheblich von der des Aminosäureausgangsgemisches. Mit anderen Worten: Die Wahrscheinlichkeit des Einbaus einer Aminosäure in ein Polymer ist nicht für alle gleich groß, vielmehr werden bestimmte Aminosäuren (z.B. die Asparaginsäure) bevorzugt. Die physikalisch-chemischen Eigenschaften der Monomeren sowie die Startsequenz eines sich bildenden Polymers bestimmen, welche weiteren Aminosäuren in die wachsenden Polypeptidketten inkorporiert werden. Angestrebt wird dabei die Ausbildung und Stabilisierung eines Zustands niedriger Energie (energiearmer Zustand) der durch zwei Kriterien bedingt wird:
Vom physikalisch-chemischen Standpunkt aus sind solche Präferenzen keineswegs ungewöhnlich, denn auch bei einer Kristallbildung lagern sich gleichartige oder aufgrund ihrer Struktur oder Ladung zueinander pasuende (komplementäre) Moleküle (oder Ionen) unter Ausschluß anderer zusammen.
Mikrosphären können durch Einlagerung zusätzlicher Proteinoidmoleküle an Größe gewinnen, überschreiten dabei aber nur selten einen Durchmesser von ~15 µm. Nach Erreichen dieses Werts teilen sie sich oder schnüren Knospen ab. Kleine Mikrosphären können miteinander verschmelzen (fusionieren). In wäßriger Lösung sind sie sehr stabil, die Stabilität sinkt jedoch beträchtlich; die Verformbarkeit steigt, sobald Lipidmoleküle in die Proteinoidschicht eingelagert werden. Diese wiederum können, wie wir schon gesehen haben, auch in der abiotischen Umwelt der Erdoberfläche vorgekommen sein.
Elektronenmikroskopische Untersuchungen von Mikrosphärenquerschnitten ergaben, daß die Proteinoide in Form einer membranähnlichen Doppelschicht organisiert (angeordnet) sind, wobei deren Dimensionen jedoch nicht mit denen der heutigen Einheitsmembran übereinstimmen.
Zusammenfassend können wir nunmehr konstatieren, daß sich aus Proteinoiden (und Proteinen) unter Mitwirkung anderer Moleküle wie Ionen, Lipiden u.a. zellähnliche Formen bilden konnten, in deren Innerem und / oder an deren Oberfläche eine Anzahl biochemischer Umsetzungen katalysiert wurden. Darüber hinaus erwarb die membranähnliche Außenschicht eine Selektivität in bezug auf Durchlässigkeit (selektive Permeabilität) bestimmter Moleküle bei Ausschluß anderer. Durch Fusion von Mikrosphären unterschiedlicher Zusammensetzung und Funktion können Fusionsprodukte entstanden sein, die die Leistungen der beiden Ausgangsformen in sich vereinigten. Gleichzeitig zeigte sich dadurch aber, daß das Auftreten, die Stabilität und die Leistungen der Mikrosphären einer Selektion unterworfen sind.
Stellen die vorgestellten Strukturen ein lebendes System dar? Die Antwort darauf ist ein klares Nein, obwohl sie über eine Vielzahl von Eigenschaften verfügen, die für lebende Zellen typisch sind: Katalyse, Kooperativität (allosterisches Verhalten), Bildung von Quartärstrukturen (self-assembly), zellähnliches Aussehen, Anordnung von Molekülen zu membranähnlichen Schichten, Teilungskompetenz, Fusionskompetenz, selektive Permeabilität, Selektion. Es fehlt aber die Fähigkeit zur identischen Reduplikation, sich also fortzuentwickeln. Das wiederum beruht auf der Unfähigkeit der Proteine, die in ihnen enthaltene Information zu speichern und weiterzugeben. Man könnte sich nunmehr einen Zusammenschluß mehrerer katalytischer Aktivitäten vorstellen, der zur Autokatalyse einer jeden der daran beteiligten Komponenten führt. Die Reaktionspartner wären dann Glieder (Elemente) eines Kreisprozesses (eines Zyklus), der sich als Ganzes replizieren könnte. Das Replikationsvermögen wäre also eine Systemeigenschaft, weil keines der Glieder alleine hierzu in der Lage ist. Die Konsequenz wäre aber auch, daß das System keinerlei Änderungen (Fehler, Mutationen) zulassen würde, da durch Mutation eines seiner Elemente der darauffolgende Katalyseschritt beeinträchtigt wäre und alle nachfolgenden Schritte unterbleiben würden. Der Zyklus würde in sich zusammenbrechen (er würde aussterben). Andererseits hätte auch die Mutante praktisch keine Chance, sich zu manifestieren, denn sie müßte geeignet sein, einen völlig neuartigen Zyklus zu initiieren; die Erfolgswahrscheinlichkeit hierzu ist beliebig gering. Wir hätten es hier deshalb auch nicht mit einer Evolution, sondern mit einer Revolution zu tun, bei der die gesamte bis zu dem Zeitpunkt angesammelte Information, also die Reihenfolge
A > B > C >........ X > A
mit einem Schlag wertlos würde und durch etwas radikal Neues abgelöst werden müßte. M. EIGEN (Max-Planck-Institut für Biophysikalische Chemie, Göttingen) hat 1971 das Zustandekommen solcher Zyklen - und deren Änderungswahrscheinlichkeiten - modellmäßig durchgerechnet und kam durch Simulation am Computer zu dem Ergebnis, daß sowohl das Alter als auch die Größe des Weltalls (nicht allein der Erde!) nicht ausreichen würden, um eine erfolgreiche Strategie auf dieser Ebene zum Zuge kommen zu lassen.
Wie kommt man aus diesem Dilemma heraus? Ist eine Entstehung des Lebens auf Nukleinsäurebasis möglich?
Nukleinsäuren können genetische Information tragen; sie haben Matrizenfunktionen, können sich daher identisch reduplizieren, und Nukleotide können unter abiotischen Verhältnissen zu Polynukleotiden polymerisiert werden. Umfangreiche Versuchsserien im Laboratorium von L. E. ORGEL ergaben, daß unter Carbodiimidzusatz Phosphodiesterbindungen zwischen Nukleotiden ausgebildet werden, wobei neben den in rezenten Nukleinsäuren enthaltenen 3' > 5'-Bindungen auch 2' > 5'- und 5' > 5'-Bindungen zustande kommen. Aufgrund der bekannten Komplementarität zwischen Purin- und Pyrimidinbasen besteht die Tendenz zur Bildung von Basenpaaren, die entweder zwischen zwei Einzelsträngen oder zwischen Abschnitten innerhalb eines Stranges beobachtet werden können. In wäßriger Lösung verknäulen sich Nukleinsäure-Einzel- und Doppelstränge statistisch; definierte Tertiärstrukturen werden nicht ausgebildet; tRNS scheint da eine Ausnahme zu sein, die jedoch auf Modifikation einzelner Basen (unter enzymatischer Mitwirkung) zurückzuführen ist.
Siehe auch:
Im Vergleich zu Einzelsträngen können Doppelstränge thermodynamischer Denaturierung (=Denaturierung, bedingt durch temperaturabhängige Molekularbewegungen) besser widerstehen. Unter Stabilitätskriterien haben sie daher einen Selektionsvorteil. Je länger ein Polynukleotidstrang ist, desto stabiler ist er. Das beruht zum einen auf der Zahl von Wasserstoffbrücken zwischen gegenüberstehenden Basen (zwei bei A = U (T) - Paaren, drei bei G = C - Paaren), zum anderen auf der sogenannten Stapelenergie (stacking energy) zwischen benachbarten Basenpaaren.
Die Fähigkeit zur Wasserstoffbrückenbildung ist nicht allein auf die o.g. Basenpaare beschränkt, denn je zwei Wasserstoffbrücken werden z.B. auch zwischen U = U -, T = T -, C = C -, A = A -, G = G - und G = T - Paaren gebildet.
Man kann sich daher fragen, warum in allen heute bekannten Nukleinsäuren nur die zuerst genannten Basenpaare vorkommen und weshalb die Nukleotide ausschließlich über 3' > 5' Phosphodiesterbindungen verknüpft sind.
Die Antwort: Polymere, bei denen stets ein Purin einem Pyrimidin gegenübersteht und bei denen die Monomeren durch stets den gleichen Bindungstyp miteinander verbunden sind, bilden regelmäßige Strukturen (einheitliche äußere Abmessung der Doppelhelix) aus. Regelmäßigkeit, und die sich daraus ergebende Komplementarität wiederum sind Ursachen einer hohen Stabilität.
DNS liegt im Gegensatz zur RNS meist als WATSON-CRICK-Doppelstrang vor. Es gibt zwar auch RNS-Doppelstränge und RNS, die abschnittsweise als Doppelstrang vorliegt, doch deren molekulare Konfigurationen sind anders. Im Unterschied zum WATSON-CRICK-Modell stehen die Basenpaare nicht senkrecht zur Achse. Die Folge ist eine meßbar geringere Stabilität gegenüber thermischer Denaturierung, und wohl deshalb hat sich DNS gegenüber RNS als Träger genetischer Information der Organismen durchgesetzt. Ein erneuter Hinweis darauf, daß Selektion nicht ein biologisches Phänomen ist, sondern als eine grundlegende Eigenart der Materie anzusehen ist.
Weshalb konnten Nukleinsäuren zu Trägern genetischer Information werden? Ein wichtiges Argument ist die Fähigkeit zur Selbstreplikation (Selbstreduplikation) aufgrund von Matrizeneigenschaften sowie ein Vorrat mehrerer Zeichen (A, T, C, G), die - formal betrachtet - in beliebiger Reihenfolge aneinandergereiht sein können, wodurch theoretisch unendlich viele Informationseinheiten zusammengesetzt werden können. Polynukleotiddoppelstränge, die nur aus C und G bestehen, sind stabiler als solche, die zusätzlich A und T enthalten, aber ihr Informationsgehalt wäre drastisch reduziert.
Eine Abfolge von Zeichen per se kann noch nicht als Information bezeichnet werden, denn zu einer Information wird nur das, was "verstanden" wird. Die Matrizeneigenschaften bilden deshalb eine Voraussetzung, um aus einer Zeichenabfolge eine Information (zur Bildung identischer, bzw. komplementärer Polynukleotidstränge) werden zu lassen. Nukleinsäuren sind folglich selbstinstruierende Informationsträger.
Bei der Replikation können durch Einbau "falscher" Basen Fehler entstehen. Ist ein solcher Fehler erst einmal eingetreten, läßt er sich nicht mehr beheben. Er wird daher in allen nachfolgenden Replikationsrunden mitgeschleppt und damit auf alle nachfolgenden Molekülgenerationen übertragen. Die Zeichenabfolge wäre dadurch verfälscht, und mit der Ansammlung weiterer Fehler kann sie vollständig verlorengehen und sukzessive durch eine andere Abfolge ersetzt werden. Erhalten bliebe nach wie vor die Eigenschaft der Selbstverdopplung.
Kann ein solches System eine Evolution durchmachen? Kann Leben auf Nukleinsäurebasis allein entstanden sein? Wie im vorangegangenen Abschnitt lautet auch hier die Antwort: Nein !
Wegen der Fehlerquote bei der Replikation fehlt den Nukleinsäuren ein Kontrollmechanismus zum Erhalt und zur Sicherung der in der Nukleotidabfolge enthaltenen Information. Es fehlt daher auch die Möglichkeit, wertvolle Information zu erhalten sowie wertvolle von wertloser zu unterscheiden. Dieser Satz impliziert, daß der Information (über das Phänomen der Selbstreplikation hinaus) ein Wert zuzuschreiben ist. Daher lautet unsere nächste Frage: Worin besteht der Zusammenhang zwischen wertvoller (genetischer) Information und dem, was wir unter einem lebenden System verstehen ?
|
|