
" I call the totality of all tissues, that are enclosed by the dermal tissue (the epidermis) and contain the vascular bundles, the ground tissue. I characterized it for the first time in the first edition of my textbook in 1868. Young and juicy organs that are covered by the epidermis alone and whose vascular bundles have up till then not been deformed by secondary growth, organs in general that have not started to develop real wood and secondary bark consist mainly of ground tissue. This may best be perceived with an apple, since all its eatable substance is made up from ground tissue alone... The most common and very likely also most original type of ground tissue is the common, thin-walled parenchyma." (J. v. SACHS).
Parenchyma or ground tissue cells are always alive, usually isodiametric and only seldom elongated. The pith of the shoots, the storage tissue of the fruits, the seeds, the roots and other underground organs are all parenchyma tissues, as is the mesophyll (the assimilation tissue of leaves). The mesophyll is reviewed separately due to its fundamental function in assimilation and its resulting importance for plant nutrition.
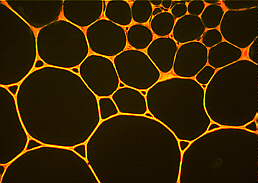
Parenchyma cells and relatively large intercellular spaces in a cross-section through the stem of Geum urbanum. The preparation has been stained with coriphosphin. The middle lamina displays a red fluorescence.
The entire function of the parenchyma seems often to be in the filling of the gaps between more specialized cells. This is expressed by the term ground tissue, which means the mass of cells into which more specialized cells, like vascular tissues or seeds are imbedded.
Parenchyma cells are not differentiated, neither morphologically nor physiologically. It is not uncommon that they contain chloroplasts. Sometimes, especially when grown without light, leucoplasts or incompletely structured chloroplasts can be found. Although hardly any cell division takes place in differentiated parenchyma, the cells keep their ability to divide: they make up the bulk of cells participating in wound healing and regeneration of plant organs. Consequently, the cells could be said to be in a kind of waiting position. They form a pool of cells that can be activated when required, for example during ontogenesis and at unusual incidents.
The meristem has already been defined as the tissue, where most cell divisions take place. The parenchyma could analogously be described as a basic tissue, whereof very different tissues develop during ontogenesis. The impact of a cell's position has already been mentioned. Whether or not a cell divides depends on its position (information of position) within the tissue. In the case of parenchyma cells, the position has a decisive influence on their further development.
Parenchyma tissues (from the pith of the shoot, for example), can be cultivated in appropriate synthetic culture media and are thus prompted to divide. They keep their capacity for cell division for several decades, if parts of the culture are transferred to fresh media at regular intervals. The cells stay in an undifferentiated state. They form aggregates, the calli (sing. callus). The cultures themselves are called tissue- or callus-cultures. The calli remain colorless, when kept in darkness, while they become usually green upon light-exposure. Addition of suitable growth hormones (phytohormones) leads to the development of shoots or roots.
The metabolism of plants produces a vast variety of substances, some of which are stored for subsequent use. In general, parenchyma cells of different organs above or underground serve as depots. The chemical features of these substances will be reviewed later. It is, at present, sufficient to know that - for cells - basic differences between small and large molecules exist. The majority of small molecules is made up by inorganic ions, organic acids and their salts, sugars, a number of nitrogen-containing compounds (like amino acids or alkaloids) and several others. Small molecules are generally stored as solutes within the vacuole. They are sometimes crystallized and these crystals are either stored within the vacuole or the plasma. Large molecules, or macromolecules, are, for example, starch and proteins. Macromolecules do rarely find a way to the central vacuole. They are stored as microscopic particles within the plasma.
Sugar and starch are the most important primary products of photosynthesis. It is commonly known that the parenchyma of fruits may contain large amounts of different sugars. The simultaneous storage of organic acids in the same cells assigns the fruits their specific, often sour-sweet taste. Starch depots can be found in seeds (within the endosperm, a parenchyma tissue), in roots, tubers (dahlia, potato) and others.
Proteins are frequently stored in seeds, usually in cells specialized for this function. A wheat grain, for example, has starch in its central endosperm cells and proteins in the form of aleuron granules at the periphery. These proteins, the storage-proteins, belong to a certain class of proteins. Their chemical composition and structure is dependent on the respective species.
When germinating, part of the macromolecules of the endosperm are degraded and used for the organization of the new plant. After mobilization of its contents, the endosperm usually dies.
Besides the already mentioned chemical compounds, parenchyma cells store huge amounts of water. This is especially striking in succulent plants, like cactus, Aloe, Agave and Aizoaceae (Mesembryanthemum family). The water content of single parts of a bamboo shoot and its content of parenchyma cells in percent can be correlated linearly.
Certain organs of plants of our regions do also contain more water than others. Among these are the flesh of fruits, onions, buds and all fleshy parts above ground.
The capacity of vacuoles for water-storage is dependent on the molarity of its solutes. It is called osmolarity.
We already got to know the vacuoles of parenchyma cells as depots for a whole range of small molecules. According to the physicochemical conditions just explained, these cells are predestined for the storage of huge amounts of water. This is the reason, why ripe fruits are always firm; the high osmotic pressure may even lead to bursting.
Most parenchyma tissues, particularly those with storage functions have only small, if any intercellular spaces at all. The thin cell walls put up hardly any resistance against the extension of the cell contents due to the high osmotic pressure (turgor) of the single cell, so that intercellular spaces cannot be formed in the first place.
The case is different in a special type of parenchymatic cells characterized by thicker walls. By splaying of such cells or tissues, air filled cavities are generated. In the shoot and the roots, these cavities are usually arranged tangentially or radially. The splaying is caused by tension differences in neighboring tissues with different growth rates. When splaying is produced by asymmetrical elongation growth, irregularly formed cells with extensive intercellular systems are created. The spongy parenchyma of foliage leaves is a typical example. Spacious aerenchymas can also be found in the pith of many monocots living in humid habitats like rush. This tissue type is also called star parenchyma because of its characteristic cell shapes.
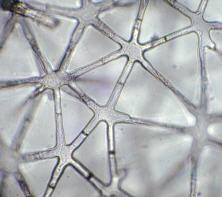
Star parenchyma with extremely large intercellular spaces in the pith of a rush stem (Juncus). This parenchyma serves as an aerenchyma (photography by W. KASPRIK).
In the submerged (growing under water) shoots of many angiosperms like the white or yellow water lily exist large air-tubes that can already be seen with the naked eye. They are separated by simple cell layers. This honeycombed construction has two effects: on one hand is a great stability achieved with only little material. This principle has also successfully been applied in technological areas. On the other hand causes the low specific weight a buoyancy that supports the floating of the shoot.
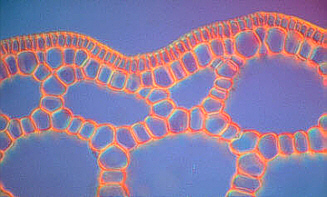
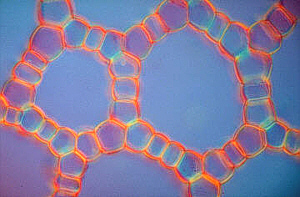
Aerenchyma cross-sections of
Hippuris vulgaris
|
|