
A special case: Monocotyledon wood is characterized by a building plan that differs completely from that of the gymnosperms or dicots. This difference results in a strongly enhanced flexibility.
Angiosperm wood in its typical structure is found almost only in dicots. First signs of wood formation occur in some monocot species, but their wood is always built inhomogenously. Contrary to the very clear architecture of gymnosperm wood, that of angiosperms is more complex and variable, partly because of a greater number of different cell types in the axial system. Most wood contains vessel members as conducting elements and, as we have seen earlier, they can be structured very differently. Over and above the wood may hold a high proportion of xylem fibres and parenchyma cells (the latter constitutes up to 23 percent of all xylem elements). The organization of rays varies also, they include almost only parenchyma cells, but parenchyma of strongly varied shaping. The differences in shape and orientation of xylem elements can be explained by the organization of the cambium.
No plant species with 'typical' angiosperm wood exists, rather can the following variables be consulted as classification and identification features for different woods:
![]() existence or non-existence of vessels
existence or non-existence of vessels
Environmental influences lead to manifold modifications, the composition of the wood changes with increasing age. Two main patterns of vessel arrangement have been recognized:
- Wood with statistically scattered pores: All vessels have about the same diameter, they are evenly distributed in every new annual ring. Examples are maple (Acer), birch (Betula) or Liriodendron, a Magnoliaceae.
- Wood with ring-shaped pores: The vessels have diameters of different size, such with large ones are normally laid out in spring. In autumn, almost only xylem fibres are laid out. This pattern is considered highly specialized. It has been shown to exist in a few species of the northern temperate zone. Chestnut (Castanea), ash-tree (Fraxinus), Robinia and certain oak species (Quercus) are examples.
Besides the clearly developed types, a number of transitional structures exist. Wood with ring-shaped lumina conducts water nearly only in the outer growth zone. In spring, the lay out of vessels is speeded up and the flow of water is ten times as high as in wood with scattered pores.
Many tropical species develop no annual rings. 75 percent of the trees of the Indian rain forest, 45 percent of those of the Amazonas Basin and 15 percent of the Malaysian trees have no annual rings. Plants of dry habitats, that are in permanent contact with the ground water like Tamarix articulata, Acacia raddiana or Acacia tortilis show steady activity throughout the year. Transplantation experiments revealed that the rhythm of cambium activity is inherited in some species, while it is chiefly controlled by extern factors in others.
The vessels are connected laterally (exchange of solutes via pits). In some species, the contact is restricted to vessels in growth zones, while it extends over several vessels in others. Ample xylem fibres are common in most angiosperm woods. In wood with ring-shaped pores, they are predominant in late wood. In some species, the xylem fibres contain living protoplasts that serve generally as starch deposits. This type of organization is estimated progressive. Lacking or small numbers of axial parenchyma cells are strikingly correlated with this type of organization.
 Rays
of angiosperm wood contain only rarely tracheids, but are constructed
from parenchyma cells, whose longitudinal axis is either orientated
radially or axially. Both single layered
and multi-layered rays occur. The rays of
dicots are often considerably larger than those of gymnosperm wood.
They range from one to many cells wide and from one to several
hundred cells high.
Rays
of angiosperm wood contain only rarely tracheids, but are constructed
from parenchyma cells, whose longitudinal axis is either orientated
radially or axially. Both single layered
and multi-layered rays occur. The rays of
dicots are often considerably larger than those of gymnosperm wood.
They range from one to many cells wide and from one to several
hundred cells high.
Just like the wood of gymnosperms, that of angiosperms is interspersed with numerous intercellular spaces, whose structure, development and content are, however, more variable. Besides channels, cavities containing resins, oils, rubber or mucigels occur often. Their walls may or may not be lined with parenchyma cells. The intercellular spaces are, just like in gymnosperms, formed by splaying or ripping. Rubber-like substances are often secreted only after degeneration of the cells. This may also lead to filling and subsequent loss of function of neighbouring cells. The production of rubber can be caused by infections, damage due to insects or physiological disorders.
Structure and arrangement of xylem elements determines the strength and other physical properties of the wood and thus its economic value. An important, though not exclusive, indication of the strength is the specific weight. The specific weight of dry, water-free wood is dependent on the mass of the cell wall material per unit volume. The weight of pure wall substance is 1.4-1.62 g/cm3; the same substance measured within the intact tissue has a weight of 0.04-1.4 g/cm3. The value 0.04 was measured for the wood of a leguminosae (Aeschynomene), while the value 1.4 has been found in a species of the Rhamnaceae family (Krugiodendron). The difference is founded on the way the tissues are constructed and on the relative amounts of the individual xylem elements.
Xylem fibres with thick walls and small lumina add to the increase of the specific weight, wide lumina and thin walls decrease it. On the other hand, the strength is amplified by xylem fibres and weakened by vessel elements. Wood with ring-shaped lumina is less resistant to some strains due to the concentration of the vessels in defined areas than wood with diffusely dispersed vessels. The famous and astonishingly stable balsa wood (Ochroma pyramidale, a species of the Bombacaceae family) has a specific weight of 0.1-0.16 g/cm2. This low weight is partly caused by the high proportion of large, thin-walled parenchyma cells.
Two general principles of construction underlie light wood. Either layers of thick- and thinwalled cells alternate or both cell types are evenly distributed and form a structurally homogeneous tissue.
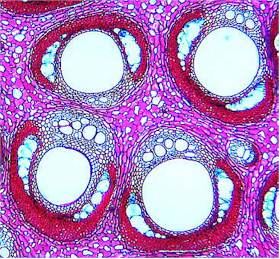
The structure of monocot wood is fundamentally different from that
of dicots. Vascular bundles strengthened by extremely tough fibres
with small lumina are embedded into voluminous parenchyma that
consists of regularly arranged cells with large lumina. The wood of
monocots is built on a 'reinforced concrete principle' that lends it
a great strength of flexibility. Just think of bamboo
or the high, slender palms that resist even
the strongest tropical storms, or the liana-like rattan
cane, whose wood is used for the production of rattan furniture.
Gymnosperm wood, however, is constructed on the 'brick
principle'. It is thus not astonishing that the wood of spruce or
pine splinters easily.
The vascular bundles of roots are enclosed by two concentric tubes of cells, the pericycle and the endodermis. The whole complex is embedded in the primary bark. Casparian strips are a typical feature of the vascular bundles of roots. Roots are able to perform secondary growth, which starts with the development of secondary cambium and leads to a restructuring of the vascular bundles. A feature of roots with secondary growth are feeder roots.
The vascular bundles of roots are constructed rather simply and show but little variations, if compared to those of the shoot. One reason could be the less variable surrounding that poses fewer demands than the surrounding above ground. Xylem- and phloem elements are combined in a central strand without pith, the stele. The root of dicots is often regarded as the basic type. The root of monocots differs only negligibly as far as the primary construction is concerned so that a separate reviewing is unnecessary. The roots of many dicot species like wood plants or perennial plants are able to perform secondary growth, while roots of monocots can, with few exceptions like Dracaena, not grown in this way. The primary cylinder of vascular bundles is as a rule composed of radially located xylem elements and peripheral phloem. The vascular elements are enclosed by two concentric tubes of parenchyma tissue, the pericycle (inner cylinder) and the endodermis (outer cylinder). The pericycle is missing in the roots of some simple aquatic plants and in some parasitic species. The whole complex is embedded into a parenchyma of large cells the primary bark.
The radial walls of the endodermis are marked by characteristic thickenings, the Casparian strips that were discovered by ROBERT CASPARI, a German botanist, in 1865/66. These thickenings contain lignin, suberin and other encrusting substances, among them sometimes also phenolic oxidation products that lend the strips a dark colour. The endodermis is regarded as a typical cell layer of roots, but it is frequently found in shoots, too, where it, just as in roots, is localized at the periphery of the vascular cylinder. In the shoots of some ferns surrounds the endodermis single vascular bundles.
The relation of the vascular bundle arrangements of roots and shoots is illustrated by the theory of steles. It is based upon the assumption that root and shoot are merely two parts of one axial unity. A continuously recurring problem is the question of the vascular bundles' reorganization in the zone of transition between both organs. It should, strictly spoken, not be talked of a root to shoot transition, since the two organs are separated by both epicotyl and hypocotyl. These are formed first in the development of the embryo and only later are shoot and roots laid out. During this early phase, vascular bundles are formed in the area of the epi- or hypocotyl that elongate, beginning at the point of initiation, simultaneously both in the direction of the shoot and the roots and in this process adopt the organ-specific arrangement.
Secondary growth occurs, as has already been mentioned above, in the roots of many dicots. In the course of this, as a rule a secondary cambium develops out of parenchyma cells that have been laid out during primary growth between the xylem rays. The cambium is supplemented by cells of the pericycle so that finally a homogeneous cambium cylinder is formed. As a result of the secondary growth, pericycle and epidermis tear apart. The consequent gaps are filled by division of additional parenchyma cells. Just like shoots, roots develop a secondary final tissue (phelloderm, phellogen, phellem).
Although the vascular bundles of roots and shoots share many principles of organization, some fundamental differences exist. The formation of the shoot's vascular bundles is normally linked to the simultaneous lay out of leaves or branches and starts at the apical meristem. Lateral roots spring from cells of the pericycle and grow from the inside towards the outside. Only in a second, independent step of differentiation is the contact of the newly developed vascular bundles to the primary (tap) root established. The lateral roots may branch again. In this way, side roots of 2., 3.,.... order develop.
Contrary to leaves, side roots are built with central symmetry in a strictly morphological sense (exceptions are, for example, the roots of Kalanchoe), while leaves and leaf stalks are normally organized dorsiventrally. But a physiological dorsiventrality seems also to exist in lateral roots (see geotropism).
It is known that roots perform two in some respects contradictory tasks. On one hand they anchor the root in the soil, on the other hand they take up water and nutrients from the soil. The anchor function demands a tension-proof root, interspersed with strengthening elements like lignification or fibres within the phloem. This function is supported by secondary growth. On the other hand lignification and formation of bark decreases the ability for water uptake and transport. Therefore has, also in roots, the 'division-of-labour-principle' been put through. Perennial plants in particular have at the endings of strongly lignified lateral or primary roots unlignified, short-lived branchings, called feeder roots. Additionally the secondary final tissues are not completely water-tight. How far they are engaged in the uptake of nutrients is not yet known.
Beside the primary and lateral roots, many mono- and dicots are equipped with so-called adventitious roots. They originate in different parts of the shoot or root. The root system of many Poaceae, for example, consists nearly entirely of adventitious roots (H. GUTTENBERG, Universität Rostock, 1940). Often are roots or parts of them including the hypocotyl used as storage organs. The parenchyma tissue of the primary bark, the xylem and the phloem swell up and numerous cell divisions take place. The newly formed deposits are secondarily supplied with vascular bundles.
|
|