
Sekundäre Pflanzenstoffe leiten sich von Endprodukten des Primärstoffwechsels ab, die durch z.T. hoch spezialisierte Biosynthesewege zu den Produkten des Sekundärstoffwechsels umgewandelt werden. Die Komplexität eines Moleküls hängt von der Zahl der Stufen im Biosyntheseprozeß ab. In kaum einer Organismengruppe sind derartige Reaktionen so vielgestaltig, und folglich auch deren Endprodukte so vielfältig, wie bei den Pflanzen.
Oft wurde behauptet, die Produkte des Sekundärstoffwechsels seien Ballast, den die Pflanzenzelle (meist in der Vakuole) speichern muß, weil sie über keine Ausscheidungsorgane für diese Substanzen verfügt. Dieses Argument ist wenig überzeugend, denn ihre Produktion ist energieaufwendig, in vielen Stoffklassen ist ein turn over nachweisbar, und wie schon dargestellt, besteht kein Zweifel darüber, daß viele der sekundären Pflanzenstoffe tatsächlich einen echten Selektionsvorteil haben. Sekundäre Stoffwechselprodukte sind in Pflanzen in unterschiedlichen Konzentrationen enthalten. Bei deren Analyse konzentriert man sich allerdings meist auf solche Substanzen, die in den betreffenden Pflanzen reichlich vorhanden sind, leicht nachweisbar sind und über deren Biosynthese - zumindest weitgehend - Klarheit herrscht.
In dieser Aussage liegt jedoch schon eine Schwäche des Verfahrens. Man betrachtet nämlich nicht alle vorhandenen Substanzen, sondern nur das, was sich als praktikabel erweist (= eine Stichprobe). Dennoch: in den letzten Jahrzehnten sind Methoden entwickelt worden, um Substanzen billig, reproduzierbar, schnell und gleichzeitig bei einer Vielzahl von Proben zu bestimmen. Es werden nur geringe Ausgangsmengen benötigt. In Einzelfällen lassen sich Substanzen auch noch in Herbarmaterial nachweisen.
Eine herausragende Bedeutung kommt dabei den chromatographischen Verfahren (Papierchromatographie, Dünnschichtchromatographie, HPLC) zur Diagnose kleiner Moleküle zu, der Gelelektrophorese zum Studium von Makromolekülen (s. folgenden Abschnitt).
Ein Vorteil der Chromatographie besteht darin, ein Substanzgemisch in einem Arbeitsgang auftrennen und die einzelnen Komponenten gleichzeitig identifizieren zu können. Dabei lassen sich nicht nur gefärbte Substanzen (oder solche, die UV absorbieren) erkennen, sondern auch jene, die durch einfache chemische Reaktionen direkt auf dem Träger (Papier, Silikatschicht usw.) anfärbbar sind.
Die Nützlichkeit der Chromatographie sei an einem Beispiel erläutert: In Farnen kommen verschiedene Flavonoide vor. Die Art Asplenium kentuckiense wurde als ein allopolyploider Bastard beschrieben.
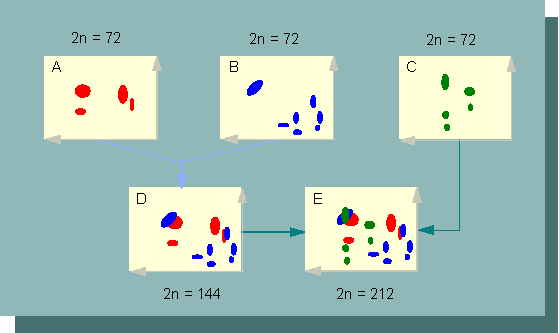
Auftrennung von Flavonoiden durch zweidimensionale Chromatographie bei drei Asplenium - Arten (Farnen), sowie ihrer allopolyploiden Bastarde . A. Asplenium rhizophyllum, B. Asplenium montanum, C. Asplenium platyneuron, D: amphidiploider Bastard: Asplenium rhizophyllum x Asplenium montanum, E. allotriploider Bastard mit den Genomen der Arten A-C (neue Artbezeichnung: Asplenium kentuckiense) (Nach D. M. SMITH und D. A. LEWIN, 1963).
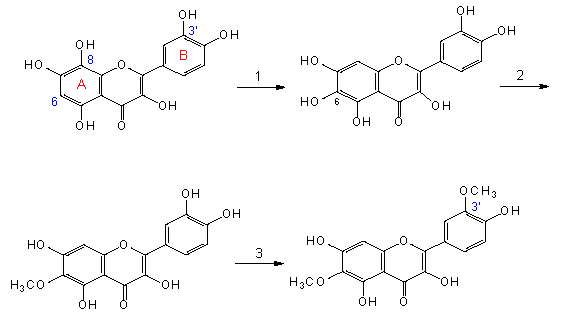
Evolution gelber Flavonole in einer Unterfamilie der Compositen. Es wird zwischen den Ringsystemen A und B unterschieden. Die Atome im Ring A werden durch einfache Ziffern gekennzeichnet, in B tragen sie einen Strich (z. B. 3´). 1. Wechsel der -OH Gruppe von 8- zu 6-Stellung (in Ring A), 2. Methylierung in 6-Stellung, 3. Methylierung in 3´-Stellung (in Ring B) (V. H. HEYWOOD et al., 1977).
Wie Karyotypanalysen zeigten, enthält sein Genom die Genome der drei Arten Asplenium rhizophyllum, Asplenium montanum und Asplenium platyneuron. Die Flavonoide der Ausgangsarten unterscheiden sich voneinander. Sie sind daher chromatographisch identifizierbar. Mehrere von ihnen ergeben ein Muster (einen fingerprint), das für die jeweilige Art, aus der das Gemisch gewonnen wurde, charakteristisch ist. Das Flavonoidmuster in dem Bastard besteht aus den Mustern der drei Ausgangsarten. Die biochemische Analyse steht damit in voller Übereinstimmung mit den cytologischen (karyologischen) Befunden.
Eine veränderte Position im Chromatogramm bedeutet Bildung eines Produkts, das der Referenzprobe ähnelt, ihr aber nicht gleicht. Zum Beispiel weiß man, daß Flavonoide in der 8-Position oder der 6-Position eine -OH-Gruppe tragen können und sich daher chromatographisch voneinander unterscheiden. Vergleichende systematische Untersuchungen ergaben, daß bei den Dikotyledonen zunächst die Tendenz zur Bildung von Flavonoiden mit der -OH-Gruppe in 8-Position entwickelt wurde, daß der Biosyntheseweg sich bei den entwickelten Taxa dann änderte, so daß wir dort vorwiegend Flavonoide mit der Gruppe in 6-Position finden.
Quantitative Unterschiede beruhen auf unterschiedlicher Effizienz der jeweiligen Biosynthesewege. Es wird nicht das Vorhandensein oder Fehlen von Genen getestet, sondern deren unterschiedliche Regulierbarkeit, oder eine unterschiedliche Regulation auf der Ebene der fertigen Protein-(Enzym)-Moleküle.
Auch die Verteilung der Stoffe in der Pflanze variiert. Das Vorkommen vieler Substanzen ist organspezifisch. Man denke dabei nur an blüten- oder fruchtspezifische Stoffe; andere wiederum findet man in einzelnen Organen oder Entwicklungsstufen in unterschiedlicher Konzentration (z.B. Unterschiede zwischen oberen und unteren, jungen und alten Blättern, im Holz, in der Rinde oder in Wurzeln).
Oft findet man in einer Population Individuen, die einen bestimmten Stoff bilden, andere, denen er fehlt. Wir haben es hier also wieder mit einem typischen polymorphen Merkmal zu tun. Neben diskontinuierlichen Unterschieden kommen auch graduelle vor, die auf Multigenwirkung oder eine komplexe Wechselwirkung zwischen Genom und Umwelteinflüssen hinweisen. (Die Chlorophyllsynthese z.B. erfolgt bei den meisten höheren Pflanzen nur nach Belichtung; doch wie Ausnahmen belegen, geht es im Prinzip auch ohne.)
Ferner gibt es populationsspezifische Muster, die mit geographischer Verbreitung oder ökologischen Ansprüchen zusammenfallen, wobei man es mit Merkmalen lokaler Populationen zu tun haben kann. Schließlich wäre noch ein letzter Einwand vorzutragen, der gegen die generelle Bedeutung sekundärer Pflanzenstoffe als Merkmal sprechen könnte. Wie an anderer Stelle dargelegt, werden in verschiedenen, meist nicht verwandten Arten (oder anderen Taxa) etliche Substanzen auf verschiedene Weise synthetisiert, obwohl am Ende gleiche oder nahezu gleiche Produkte entstehen.
Es gilt dabei allerdings die Regel, daß die Wahrscheinlichkeit, mit der eine Substanz auf unterschiedlichen Wegen gebildet wird, mit zunehmender Komplexität ihrer chemischen Struktur abnimmt. Sie gilt jedoch nicht für Substanzen, deren Biosynthese (vom Endprodukt des Primärstoffwechsels ab) nur einen Schritt erfordert. So findet man z.B. Nicotin in der Gattung Nicotiana und in Schachtelhalmen (Equisetum). Coffein tritt in vielen, z.T. nicht verwandten Angiospermenfamilien auf. Andererseits gibt es eine ganze Reihe von Substanzen, die für bestimmte Pflanzengruppen typisch sind. Daher wurde auch der Vorschlag unterbreitet, sie für die Großgliederung der Gymno- und Angiospermengruppen heranzuziehen.
So enthalten z.B. die meisten Coniferen der nördlichen Hemisphäre Terpene; Alkaloide sind bei den Ranunculaceen verbreitet, Tannine bei Fagaceen usw. Einige Substanzen sind weit verbreitet, fehlen aber in einzelnen Taxa; beispielsweise fehlt die Ellagsäure bei den meisten Gattungen einer Unterfamilie der Rosaceae. Andere Substanzen sind für nur eine Familie, Gattung oder Art charakteristisch. So kommt die Aminosäure Canavanin nur bei den Leguminosen und das Monosaccharid Acofriose nur in Acokanthera friesiorum (Familie Apocynaceae) vor.
In einigen Familien, z.B. den Euphorbiaceae, den Apocynaceae und den Asclepiadaceae, gibt es komplex strukturierte, latexhaltige "flüssige Gewebe". Ein ähnliches System kennzeichnet eine Tribus der Compositae: die Cichorieae; den anderen Triben fehlt es, statt dessen findet man dort oft aromatische Harze. Auffallend ist die alternative Ausprägung der Blütenfarbstoffe: Anthocyane einerseits, Betalaine (Betcyanin, Betaxanthin) andererseits. Letztere sind auf die meisten Familien einer Gruppe mit der alten Bezeichnung Centrospermae beschränkt. Wie die Listen in der Tabelle zeigen, müßte man nach diesem Merkmal die Cactaceae den Centrospermae zuordnen, die Molluginaceae und die Caryophyllaceae aber ausschließen. Hier führen morphologische Daten und Biochemie der Blütenfarbstoffe also zu unterschiedlicher Bewertung. Durch elektronenmikroskopische Untersuchungen zeigten H.-D. BEHNKE und B. L. TURNER (1971), daß sich die beiden zuletzt genannten Familien auch durch strukturelle Unterschiede ihrer Siebelementplastiden von den übrigen Centrospermae unterscheiden. Dieser Befund stützt die durch Blütenfarbstoffanalyse gewonnene Klassifikation. Demnach hat man die Centrospermae (=Unterklasse Caryophyllidae) in die Ordnungen Caryophyllales und Chenopodiales aufgeteilt.
Die Analyse sekundärer Pflanzenstoffe eignet sich auch, um eine geographische Verbreitung von Pflanzenarten sowie den Diversifikationprozeß; nachzuvollziehen. In der im tropischen Südamerika verbreiteten Lauraceengattung Aniba kommen artspezifisch alternativ entweder Neolignane oder Pyrone vor. Beide Stoffklassen werden über den Shikimatweg gebildet; im zweiten Teil der Biosynthesekette trennen sich die Wege, und nur einer kann eingeschlagen werden. Die Verbreitungsgebiete der einzelnen Arten weisen darauf hin, daß Arten mit gleichen Endprodukten allopatrisch oder vikariierend sind. Sympatrisch sind nur solche Aniba-Arten, die sich in bezug auf die obengenannten Stoffe unterscheiden.
In Gentianaceen kommen Xanthone vor. Das Xanthonringsystem ist bei europäischen Arten einfach gebaut, in nordamerikanischen ist der Ring A modifiziert, in asiatischen der Ring B. Dies wiederum legt die Annahme nahe, daß die Gentianaceen in Europa ihren Ursprung hatten und sich von dort aus unabhängig in Richtung Asien und Nordamerika ausgebreitet haben (O. R. GOTTLIEB und K. KUBITZKI, 1983).
Im Verlauf der Evolution nimmt in der Regel die Komplexität auf verschiedenen Ebenen zu. Nachdem ein Maximum erreicht ist, kommt es oft wieder zum Abbau, bzw. zu einer Vereinfachung oder Verkürzung von Reaktionswegen. So war die Bildung von Holz eine Voraussetzung zur Evolution großer Landpflanzen. Bäume findet man unter den Pteridophyten, dann sogar ausschließlich in der Gruppe der Gymnospermen, und schließlich in vielen, vor allem primitiven Angiospermenfamilien. Im Verlauf der Diversifikation der Angiospermen trat sekundäres Dickenwachstum mehr und mehr in den Hintergrund. Kräuter gewannen in vielen Biotopen die Überhand, und vielfach eroberten sie Standorte, die für Bäume verschlossen waren. Mit zunehmender Vervollkommnung dieser Entwicklung ging die Verkürzung der Biosynthesewege bestimmter sekundärer Pflanzenstoffe einher. Die Ausgangsstoffe für das nur noch in geringer Menge benötigte Lignin konnten zur Produktion neuer Substanzen eingesetzt werden.
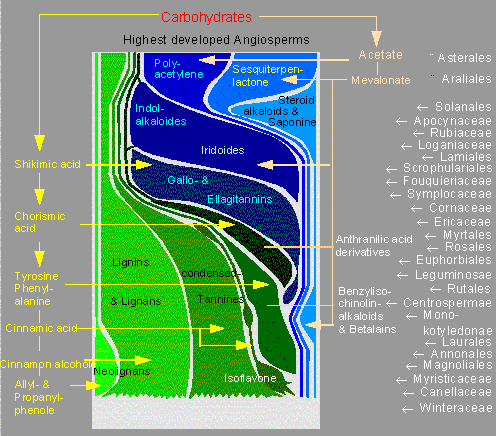
Verkürzung von Biosynthesewegen im Verlauf der Angiospermenevolution. Verteilung einiger biogenetischer Gruppen von sekundären Pflanzenstoffen mit allelochemischer Wirkung bei den Angiospermen. Das Schema zeigt, daß Derivate des Shikimat-Weges (in Grüntönen dargestellt) zunehmend von solchen des Mevalonat- und Acetat-Weges (in Blautönen dargestellt) ersetzt werden: Entstehung krautiger Formen. Die Beteiligung von Acetat bei kondensierten Gerbstoffen und Isoflavonen ist nicht berücksichtigt. Die angegebenen Familien und Ordnungen sind Beispiele für die jeweiligen Konstellationen von Pflanzenstoffen. Ein lineares Abstammungsverhältnis zwischen ihnen wird nicht angenommen (Nach K. KUBITZKI und O. R. GOTTLIEB, 1984).
Einerseits sparte die Pflanze damit Energie ein, ein ganz wesentlicher Punkt, um von perennierender zu annueller Lebensweise überzugehen, andererseits erweiterte sich das Spektrum der sekundären Pflanzenstoffe. Viele dieser Stoffe sind toxisch, sie sind als Abwehrstoffe gegen Insekten, andere Invertebraten und Vertebraten herausgezüchtet worden (E. STAHL, 1888). Die Vielfalt der Stoffe ermöglicht eine Entstehung zahlreicher Abwehrstrategien, das wiederum ist vorteilhaft, denn gegen eine einzige Abwehrstrategie könnten die Tiere ihrerseits leicht Mechanismen entwickeln, um sie außer Kraft zu setzen.
Wie die vorstehenden Beispiele zeigten, ist die Chemotaxonomie ein geeignetes Hilfsmittel zur Klärung vieler Einzelfragen. Man kann aber nicht sagen, sie sei dem konventionellen Vorgehen (Vergleich und Auswertung morphologischer Merkmale) überlegen und in der Lage, alle offenen Probleme zu klären.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de