
Since the beginning of the century has it been known that unstable or variable gene loci occur in plants although the drastically enhanced mutability and the increased back mutation rate could not be explained at first. The decisive breakthrough was accomplished by B. McCLINTOCK with her studies on maize chromosomes published in 1947 and 1951. The basis of these were her earlier observations and analyses (1938) on breakage-fusion (reunion)-bridges. Their occurrence could be correlated with the restructuring in chromosomes
Bridges are formed during anaphase whenever two chromosomes fuse at their ends generating a fusion product with two centromers. If these two are subsequently torn to different poles then will inevitably occur a chromosomal fraction. During the following S-phase of the interphase nucleus is a chromatid with a fraction at its terminus replicated in just the same way as the other chromosomes leading again to a fusion of the homologous chromatids. Consequently can a chromosome consisting of just one chromatid but two centromers be found in the subsequent mitosis instead of a chromosome out of two chromatids and one centromer. The consequence is a second fraction during anaphase where the second round of the cycle starts.
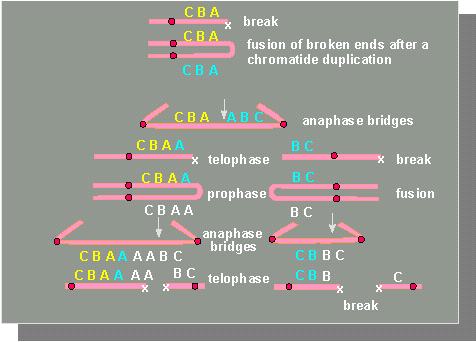 B. McCLINTOCK recognized that the fraction cannot occur at any
site of the chromosome but is restricted to certain sections that
she called Ds (dissociation). These were obviously DNA
segments contributing to the formation of translocations, deletions,
inversions and to the generation of ring-shaped chromosomes.
B. McCLINTOCK recognized that the fraction cannot occur at any
site of the chromosome but is restricted to certain sections that
she called Ds (dissociation). These were obviously DNA
segments contributing to the formation of translocations, deletions,
inversions and to the generation of ring-shaped chromosomes.
The first fraction causes similar fractions in the mitosis cycles of following generations. They happen during ontogenesis at different times and sites.
The segment Ds, a mutator gene, behaves like a multiple allele (or, even better, like a pseudoallele) that can be located at different gene loci. It may also vary in structure. This mutator can insert itself into other genes thus rendering them inactive. It is a control element that changes its place within the chromosome, jumping or wandering around and causing mutations wherever it inserts (the mutators are also called jumping genes).
It soon became clear that a further set of elements has to exist: the Ac (activation) elements. A chromosomal fraction or a translocation of a Ds element has to be supported by an Ac element. An Ac element can also be regarded as a multiple allele. It may occur at the most different sites in all chromosomes. To analyze its effect further concentrated B. McCLINTOCK on the study of genes that determine the colour of maize grains.
One of the most important is the C-locus that causes a dark red staining of the aleuron layer and the pericarp of the maize grain in a dominant condition. If a Ds-element jumps into the gene, colour synthesis is interrupted and colourless (yellow) grains result. An Ac activity within these grains causes a pattern of dark red areas on a light ground. This is explained by a reestablishment of the old state since the Ac element removes the Ds from the C-locus. This happens in several cells during the development of the maize grain. These cells are the origin of the aleuron layer and the pericarp and the back mutation can only be perceived in the clones that form out of the changed cells.
 Today are a number of gene loci known that can be influenced by
the Ds-Ac-system or other control elements. The detection of the
spm-system (suppressor-mutator) and the elucidation of its function
showed that the control elements do not only act as switches (a
yes/ no decision) but that they do modulate the degree of gene
expression, too.
Today are a number of gene loci known that can be influenced by
the Ds-Ac-system or other control elements. The detection of the
spm-system (suppressor-mutator) and the elucidation of its function
showed that the control elements do not only act as switches (a
yes/ no decision) but that they do modulate the degree of gene
expression, too.
The genetic analyses of B. McCLINTOCK were not understood for years. Only when insertion elements and transposons were found in bacterial DNA during the late sixties did an analogy between them and the control elements show up. These genetic data fitted neatly with molecular biological models (B. NEVERS and H. SAEDLER, 1977, H-P. DÖRING and P. STARLINGER, 1984).
Mrs BARBARA McCLINTOCK was awarded the Nobel prize for medicine and physiology for her pioneer achievements.
P.NEVERS, N. S. SHEPHERD and H. SAEDLER listed the 'unstable plant genes' described in literature at the beginning of 1986. It shows that such genes have been found in more than 30 species. Many of the respective mutants with names like variegate, marmorata, maculata or variabilis are on the market as ornamental plants due to their irregularly spotted flowers or leaves.
A transposon is a piece of DNA that is flanked by two insertion
elements (IS elements) oriented opposite (a palindrome).
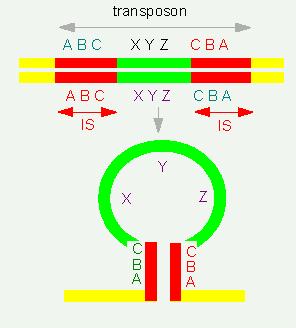
Structure of a Transposon: At the top: within a DNA double strand, at the bottom: when denaturing the double strand (just one of them is shown). The inversely oriented IS elements (insertion elements) at the ends of the transposon form double stranded segments.
The IS elements exert an influence on the expression of neighbouring genes. They are able to induce deletions and they have the ability to integrate and disintegrate into the bacterial genome. During these processes can genes on a transposon that are flanked by IS elements also be transported. Integration and disintegration, respectively, occur only, if the cell supplies the necessary enzymes. This means that two components are needed for all structural changes of the DNA and the chromosomes (deletions, duplications, inversions, insertions, etc.): the enzymes and certain recognition sequences within the DNA: the transposons. This, too, means that these processes can occur at numerous sites of the DNA but not at any site (site-specific recombination). Complementary nucleotide sequences take part in the recognition process that are distributed over the genome. The exchange of genetic information occurs via a recombination process. But contrary to the processes during meiosis where homologous chromosomes pair over their whole length are only relatively short sequences paired here. It is therefore also spoken of illegitimate crossing-over.
The analogy of McCLINTOCK's control elements and bacterial transposons
was proven by molecular biological analysis (isolation, cloning
of the DNA sections in bacteria and subsequent sequencing).
The transposons of maize occur often in pairs. One of the elements is always autonomous, i.e. it can exert all its functions by itself while the other is passive and can only be activated when the active element is present. This way of functioning has already been discussed when talking about the Ac-Ds pair.
Similar conditions can be found in Drosophila. It showed that the active element here differs from the passive only in an intern deletion. Consequently were also the Ds- and Ac sequences examined. N. FEDEROFF (Carnegie Institution of Washington) found out when hybridizing isolated Ds with Ac DNA that the first differs actually from the latter in that it lacks an intern segment of roughly 200 base pairs. The analyzed Ac element consisted of 4300 base pairs, the respective Ds element of 4100. Another Ds allele had just 1700 base pairs.
Quite a number of Ds elements with differing lengths have been isolated until now. The shortest (Ds1) has only 405 base pairs (isolated by J. C. OSTERMANN and D. SCHWARTZ in 1981). P. STARLINGER and his collaborators (Genetic Institute of the University of Cologne) isolated an insertion of more than 30,000 base pairs flanked by Ds elements on both ends out of mutants with a defect sucrose synthethase gene [the Sh- (shrunken) locus].
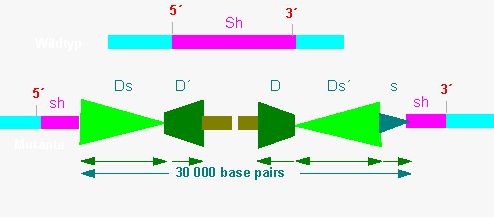
Insertion within the shrunken locus of maize. The gene encodes a sucrose synthethase (one of the most important proteins within the endosperm). In one of the mutants has it been inactivated by the insertion of a 30,000 base pair long segment. This segment is flanked by Ds elements of different orientation. Since the ends of the whole segment contain complementary sequences (< >) are they able to pair (supported by an enzyme called transponase). They form a transposon structure and can be excised from the sucrose synthase gene again. The excision may , too, take place at another site as long as complementary sequences are present (< > or > <). The model shows that the fragment between the Ds elements is unimportant for the excision (and the subsequent insertion at another site). Even a complete gene can be transferred as long as it contains Ds elements on both ends (according to P. STARLINGER, 1985)
The finding that each element contains either two complete or one complete and one incomplete Ds element in opposite orientations is quite remarkable. The sequences at the ends of Ds elements are always
T-A-G-G-A-T-G-A-A.
In Ac elements may the terminal T be replaced by a C. Two of the Ac elements sequenced in Cologne are 4563 base pairs long.
It has already been said that control elements are not solely restricted to maize. H. SAEDLER and his collaborators (Max-Planck-Institut für Züchtungsforschung, Cologne) analyzed their molecular structures in Antirrhinum majus, others studied petunias. The observation that control elements that behave like transposons exist in the plant kingdom offers a credible (and potentially provable) explanation for a host of unexplainable or hardly noticed phenomenons. Pattern formation like in the irregular colour distribution of flowers or in the uneven distribution of chlorophyll-containing and chlorophyll-free areas of a leaf are the visible expression of changes of gene activity during ontogenesis by leaps and bounds. These changes do not follow a rigid, reproducible development scheme like in sectorial or periclinal chimeras, but they occur statistically. The assumption that control elements or control element systems (including the enzymes necessary for the transfer) are at work here offers a consistent explanation. Beside their meaning for the expression of characteristics during ontogenesis do they presumably also play an important part in the speeding up of evolutionary processes. As we will discuss later, do the genomes of related species often differ in their degree of ploidy, in restructured chromosomes, in a reduction or multiplication of their heterochromatin or in the loss of activity of single genes. This leads to the assumption that the evolution of many-celled, eucaryotic organisms is not that much based on the accumulation of point mutations but more on the restructuring of existing genetic information. Our knowledge about control elements/ transposons shows how these changes (can) take place on a molecular level.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de