Algae are the plants with the simplest organization. Many of them are single-celled, some have no cell wall, others do though its composition and structure differ strongly from that of higher plants. They are good specimen for tracing back the evolution of the cell wall. Primitive cell walls do not fulfil the same requirements as that of higher plants.
It seems quite likely that a structure like that of the cell wall has developed several times in the course of evolution. All archaebacteria, eubacteria and blue-green algae (cyanobacteria or blue-green algae) have complex walls with an energetically rather costly biosynthesis. Neither in composition nor in biosynthesis do they have any common ground with the cell walls of plants.
Although the evolution of plants from early eucaryotic cells is not known in detail, is it commonly agreed on that primitive algae are flagellates closely related to the non-green flagellates. Many, though not all species of this stage of evolution, among which the euglenophyta are typical green representatives, have no cell wall. It is not only a simple membrane, but by a pellicle of already quite complex organization, that separates them from the surrounding. It consists mainly of glycoproteins organized in regular patterns the way two-dimensional crystals are. Helical ribs wind round the cell's surface.
Most single-celled algae like the Volvocales possess real cell walls. The most-studied species is Chlamydomonas reinhardii. Its wall lacks long, fibrillary carbohydrates. Most of it is made up by glycoproteins, and even here can an extensin-like protein rich in hydroxyproline be found. Among the identified sugar residues are arabinosyl-, galactosyl- and mannosyl residues. In the electron microscope does it seem as if the wall consisted of seven layers. The middle layer contains an extensive grid-shaped framework of polygonal plates consisting mainly of the mentioned glycoproteins, while the layers above and below display fibre-like structures. The thickness of the outer layer varies since it includes components that the cell takes up from its surrounding.
This indicates a main function of the cell wall of simple, single-celled algae: it mediates between the cell and its surrounding. It protects not only the cell but serves, too, communication with cells of the same or other types. It has to be permeable for metabolites and regulators and / or to carry receptor molecules with which it may contact other cells. The diversity of these functions (and their specificity) caused the evolution of a variety of differently structured cell walls.
In many-celled plants is the communication via the whole cell surface largely restricted. Contact with neighbouring cells develops in the course of tissue formation. Strength is in this respect a decisive and limiting criteria. The exchange of compounds between cells occurs via specific openings in the wall (pits, plasmodesmata). The functions originally performed by one structure are now distributed onto two different structures.
The main structural elements of all plant cells are polysaccharides. Differences in their chemical composition cause fundamentally different physical properties. No plant cell wall consists only of one class of molecules. The interactions of the different molecules produce properties that allow to distinguish the cell walls of certain classes.
In many classes of algae is cellulose already the main structural element of the wall, though remarkable variations of the fibrillary structure exist. Reliable X-ray analytical data prove that is mostly crystalline in cells of algae, too. Differences in the type of the flexor reflexes hint at the fact that cellulose could aggregate in many more or less uniform crystalline structures. Such reflexes are a measure for periodic distances at the molecular level, which may differ considerably from species to species and are specially large in Rhodophyta.
In some classes of algae exist only disperse textures, while others (specially many Chlorophyta-species) have a higher degree of organization (layers of parallel microfibrils). Such layers do usually alternate with layers of an amorphous material. No clear difference between primary and secondary cell wall exists in most algae. Where such a distinction is possible, differ the causes usually from that in higher plants.
Mannanes In a number of marine green algae (Codium, Dasycladus, Acetabularia, etc.) as well as in the walls of some red algae (Porphyra, Bangia) constitute mannanes the main structural elements. They are linear and the mannosyl residues are 1 > 4 glycosidically linked. Hydrogen bonds that are (just like in cellulose) the cause of the partially crystalline organization of microfibrils may develop. In Codium the carbohydrates are tightly associated with protein
Xylanes are polymers where the beta-D-xylosyl residues are linked via 1 > 3 and 1 > 4 glycosidic bonds. In contrast to the polymers discussed until now, are xylans partially ramified. In species with xylan-containing walls exists nevertheless a layered structure and an orientation of the microfilaments. They contain mostly linear polymers.
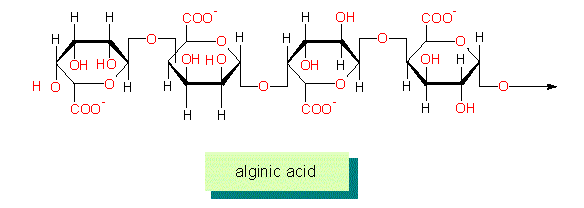
Alginic Acid: alginic acid and its salts, the alginates are important components of the walls of phaeophyta (brown algae). They are singular in many respects. They consist exclusively of uronic acids: mannuronic acid and beta-L-glucuronic acid in changing ratios and of small amounts of beta-D-glucuronic acid.
Beside homopolymers exist also heteropolymers in many algal groups, partially exist species specific differences, an indicator of the fact that the single species contain different sets of enzymes.
The alginates of brown algae exist both within the cell wall and in the intercellular substance. Their part in the cell wall may be as high as 40 per cent of the dry matter. They have a high affinity for divalent cations (calcium, strontium, barium, magnesium) and the tendency to gel. The main portion of the magnesium ions isolated from brown algae stem from the alginic acid fraction.
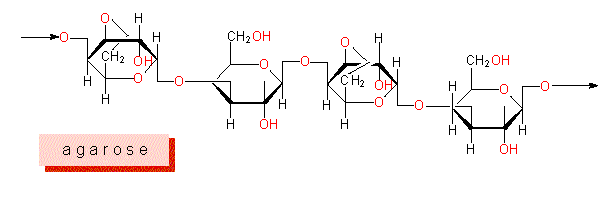
Sulfonated Polysaccharides: polysaccharides: polysaccharides whose monomers are esterized to sulfuric acid residues and are moreover partially methylated have been detected in nearly all marine algae. They occur partially in the cell wall itself and partially in the intercellular substance. Sulfonated galactanes are typical for many red algae, depending on their origin are they called agarose, carrageenan, porphyran, furcelleran and funoran.
L- and D-galactose, which are linked by beta 1 > 3 or alpha 1 > 4 glycosidic bonds form the basic pattern of agarose and porphyran, in the latter alternate L- and D-galactosyl residues. Carrageenan and furcelleran contain exclusively D-compounds. Just like in alginates is the formation of gelatine one of the most important physical properties of this family of molecules. Agar, whose basic unit is agarose, is yielded mainly from Gelidium and Gracillaria, both genera of red algae.
The extraordinary binding types of agarose and carrageenan lead to specific tertiary structures.
Further Cell Wall Compounds. A number of algae contain mineral cell wall components. Silicon, for example, is the main component of the diatom shell, though it occurs also in the cell walls of other groups of algae. Silicon-containing scales enclosed the chrysophyt Synura. In some brown algae and in the green algae Hydrodictyon is silicon a cell wall component. Diatoms take silicon up as silicate. The process is dependent on oxygen and temperature, it consumes energy and it is dependent on the presence of divalent sulphur.
Sporopollenin is an isoprene derivative. It is a component of pollen cell walls, but was also detected in the walls of some green algae (Chlorella, Scenedesmus, etc.).
Calcium: calcium encrustations of cell walls have on several occasions be described. They seem to be especially common in species of tropical, marine waters. Some species participate in reef formation. Calcium is always deposited as calcium carbonate. Calcium carbonate occurs in two different crystalline states: calcite and argonite. Calcite is produced in the walls of some groups of red algae and in charophycea, while argonite is produced by some green (Acetabularia, etc.), brown and red algae. Both states do not occur simultaneously in one species.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de