
Max-Planck-Institut für Entwicklungsbiologie, Spemannstr. 35, D-72076 Tübingen, Germany
- The activator-inhibitor reaction
- Formation of periodic structures
- Activation of genes under the influence of morphogenetic signalling
- Formation of filament-like branching structures
- Pattern formation in secondary embryonic fields:
differentiation borders obtain organizing properties
- Conclusion
- References
The problem of pattern formation
The egg cannot contain the final pattern
Organizing regions and the concept of positional information
Generation of a primary pattern by autocatalysis and lateral inhibition
The generation of the complex structure of a higher organism within each life cycle is one of the most fascinating aspects of biology. The similarity of identical twins underlines of how reproducible this process can be when the genetic information is the same. Reference to the genes, however, does not provide on its own an explanation for the generation of spatial structures since with each cell division both daughter cells obtain, as the rule, the same genetic information. This leads to the question of how different parts of the developing organism can become different from each other. How can patterns emerge in an initially more or less structure-less system? How can different genetic information be activated at different positions?
Basic concepts, such as Positional Information (Wolpert, 1969) or the embryonic organizer (Spemann and Mangold, 1924) have been derived from experiments involving perturbations of normal development. From the observed regulatory phenomena one cannot directly deduce the molecular mechanism on which development is based. We have used such observations to develop specific models for different developmental situations. By computer simulation we have shown that the regulatory features of the models correspond closely to the experimental observations. More recently, the new tools of molecular biology have opened a second inroad. In this way, several of these models have obtained direct support. In the present article, an overview of some of these models is given. Detailed mathematical equations and software that run the simulations on a PC can be found in my book "The algorithmic beauty of sea shells" Springer Verlag, 1998.
Although many eggs have pronounced asymmetries, the final pattern cannot already be present in the egg in a hidden form. At early stages, many embryos can be fragmented into two parts and each part forms a complete organism. The sea urchin embryo at the 16 cell stage is an example. Historically the first organisms in which the an experiment of this type has been performed, is the small freshwater polyp Hydra (Fig. 1). In some vertebrates, this can also occur spontaneously, giving rise to identical twins. This indicates that in these cases the early embryo is not a mosaic-like arrangement of differently determined cells that have a fixed fate. Instead, a communication must exist between different parts, such that the removal of some parts become detected and the missing parts replaced.
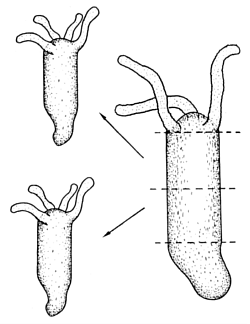
Fig. 1: Regeneration: Fragments of a Hydra regenerate the complete animal. In this process, the polarity is maintained: new head appears always at the side pointing towards the original head.
Some small specialized regions obviously play a decisive role for the overall organization for the development of an organism. Such organizing regions direct pattern formation in the surrounding tissue. An example of such a region is the dorsal lip of the amphibian embryo (Spemann and Mangold, 1924). After transplantation to the opposite site, a secondary embryonic axis can be induced. Similarly, small pieces of near head tissue of a Hydra, when transplanted into the body column of another animal, can induce there a complete head with tentacles. Small regions with an organizing influence on the surrounding tissue are also found in the generation of substructures. For instance, at the posterior margin of a chick wing bud a small nest of cells exists that organizes the anteroposterior pattern of the limb. Transplantation of these cells into a more anterior position causes mirror image duplications of the bones of the wing with a "little finger" at the anterior and at the posterior wing margin (Tickle et al., 1975).
To account for the long range effect of small specialized regions and for the spatial continuity observed after many experimental interference, the "positional information" scheme has been proposed by Wolpert (1969). Organizing regions are assumed to be the source of morphogenetic substances. By diffusion and decay graded distributions of such substances are generated. The cells are assumed to interpret this positional information by a concentration-dependent gene activation. The local concentration is a measure for the distance from the organizing region. After transplantation of such a source into an ectopic region, the surrounding tissue becomes exposed to the graded concentration profile of the morphogen. When competent cells are exposed, they follow the same pathway as the tissue that normally surrounds the organizing region. Meanwhile clear cases of morphogenetic gradients have been found. In the Drosophila egg, a localized deposition of messenger RNA of the bicoid gene leads to a graded distribution of the corresponding protein (Driever and Nüsslein-Volhard, 1988, 1989).
The assumption of a localized source, however, only shifts the problem. The question remains of how a localized source can emerge in an initially more or less homogeneous tissue or at a particular position within the developing organism. As shown in Fig. 1, complete pattern regulation can be possible, indicating that pattern formation can be a highly dynamic process.
Pattern formation is not a privilege of living systems. High sand dunes are formed although the sand is permanently redistributed by the blowing wind. Sharply contoured rivers are formed by erosion although the rain is more or less homogeneously distributed over a particular region. The formation of a lightning from a diffuse cloud is a further example. Together with Alfred Gierer I have shown that the crucial condition for pattern formation is local self-enhancement and long range inhibition (Gierer and Meinhardt, 1972; Gierer, 1981; Meinhardt, 1982). It is easy to see that the examples mentioned above, sand dunes, rivers and lightning, are also based on this mechanism. A small elevation above a homogeneous steady state has a strong feedback on its further increase such that the deviation becomes more and more pronounced. This increase becomes eventually restricted by an antagonistic effect, resulting either from a depletion of a prerequisite derived from the surroundings or by an inhibition that spreads out from the self-activating centre.