
Stomata are functional units of the epidermis serving the exchange of gases between the intercellular spaces of the plant and its surrounding. They are especially common – and of characteristic shape – at the epidermis of the leaf’s underside of most species. Their development differs from plant group to plant group, but unequal cell divisions are always involved. A functional unit consists of the guard cells themselves that contain nearly always chloroplasts, and of their neighbouring subsidiary cells that are usually devoid of chloroplasts.
It is well-known that stomata open in a humid surrounding, and close when it is dry. J. HEDWIG was the first researcher of the 18th century who understood that they work as ‘transpiration openings’. Today, it is spoken of their involvement in transpiration instead. It was thus not far-fetched to put the stomatas’ mechanism back to changes of the turgor, and to classify them as turgor movements. This assumption was verified in plant-anatomical studies of the stomata (and especially of their cross-sections). The outer and the inner walls of many species are strongly thickened compared to the lateral walls. They put therefore up considerable esistance to every deformation. The wall between guard and subsidiary cell and the wall that makes up the opening are rather thin and are easily stretched. This asymmetry of the cell’s structure and of the wall thicknesses explains the directed movement caused by the turgor. The mechanics of the guard cells becomes thus understandable. The explanation how the movement is elicited and regulated was more difficult to find. Water proved to be an important, though not the only controlling factor of guard cell movements.
![]() The concentration of carbon dioxide,,
The concentration of carbon dioxide,,
![]() light,,
light,,
![]() potassium-ions, and
potassium-ions, and
![]() abscisic acid (ABA) were shown to be as important.
abscisic acid (ABA) were shown to be as important.
Guard cells can emit water into three different directions:
- outwards,
- into the neighbouring subsidiary cell, and
- into the respiratory cavity that is a part of the intercellular system lying beneath the guard cells.
An equilibrium between the water vapour of the atmosphere and the respiratory cavity results when the stomata are opened. Plants form an intermediate distributor, since a large difference in water potential between the moist soil and the normally dry atmosphere is very common. Plants profit from the concentration gradient (that is a gain of energy for them), while the closing movements of the stomata exert a decisive, regulating influence. They close when too much water is lost, or when not enough supply exists. The osmotic pressure of the stomata is far larger in the guard cells than in the subsidiary cells. This ratio shifts in favour of the subsidiary cells when the stomata are closed.
The stomata of most plant species are closed in darkness. Light stimulates opening. The action spectrum is similar to that of photosynthesis. Blue light is especially effective. The stomata of CAM-plants, like Crassulaceans, are opened during the night. They depend on the accumulation of carbon dioxide during the night. These plants store the carbon dioxide as malate or aspartate and feed it into the CALVIN cycle during daytime. Opened stomata would cause intolerable transpiration losses in the areas that CAM-plants live in.
A low concentration of carbon dioxide (in the respiratory cavity) causes the stomata to open, a high concentration leads to their closing. Photosynthesis starts with the first light of the day, because enough carbon dioxide has been accumulated. Photosynthesis takes place in guard cells, too, since they contain chloroplasts – in contrast to the subsidiary cells. This activity again is related to the rise of the osmotic value and thus also to the opening of the stomata.
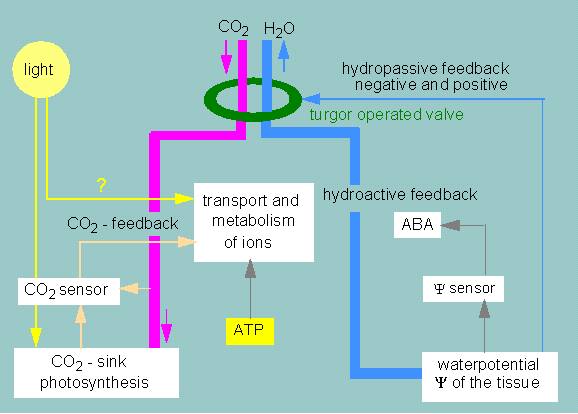
Controlling cycles regulating the movements of stomata. A guard cell is depicted schematically (according to K. RASCHKE, 1975)
That much about observations and measurements. But on what is the rise or lowering of the osmotic value based? Good evidence exists that light exerts its effect mainly by decreasing the intercellular and intracellular concentration of carbon dioxide. Carbon dioxide is faster consumed than supplied due to the mesophyll’s photosynthesis. This causes an intercellular deficit close to the stomata’s opening. The photosynthetic activity within the guard cells themselves leads to a decrease of the intracellular level of carbon dioxide, and causes simultaneously that water is drawn from the subsidiary cells.
It turned out that the water-uptake is preceded by an uptake of potassium ions. Potassium ions are actively pumped (by a potassium pump) from the subsidiary cells into the vacuoles of the guard cells. At the same time, anions (chloride, malate) accumulate within the vacuoles. Protons are given off to the subsidiary cells. The ion flows are quantitatively enough to explain a rise of the turgor that is large enough for the guard cell movements, but the primary stimulus is still not clarified. The following questions remain to be answered:
![]() What is the reason for the potassium pump’s sudden rise in activity?
What is the reason for the potassium pump’s sudden rise in activity?
![]() Is the flow of protons a result or a cause of the potassium ion transport?
Is the flow of protons a result or a cause of the potassium ion transport?
The actual importance of the potassium pump for the guard cell movement is best demonstrated with a fungal toxin called Fusicoccin (from the fungus Fusicoccum amygdali). This toxin activates the potassium pump. If consequently the toxin is applied to the stomata, then the loss of water becomes higher than its supply resulting in withering. The biological advantage for the fungus lies in the open stomata since they are, beside wounds, the only places where its hyphes can penetrate the leaf tissue. The total water balance is hardly or not at all influenced, as long as the effect of Fusicoccin remains a local one. In principle, one open stomata is enough for a hype.
The plant starts an enhanced production of abscisic acid in case of a water shortage. The abscisic acid is transported to the guard cells, where it is stored. It inhibits the potassium pump, hinders the production of an osmotic pressure, and does thus cause the closing of the stomata.
Conclusively, we can say that the opening and closing is regulated by two independent controlling cycles (that of water and that of carbon dioxide). Regulation via water potential is an effective mechanism. On one hand, the amount of water in the direct vicinity of the guard cells is calculated, and on the other hand, the water potential of far-away parts of the tissue is computed via the effect of ABA. The water and the carbon dioxide cycle may compete in case of closed stomata, since carbon dioxide is usually a limiting factor in photosynthetically active tissues. The stomata remain nevertheless closed at simultaneous lack of water. The rate of photosynthesis decreases to a low level, though it does not stopped at all due to the carbon dioxide that is repeatedly produced anew in considerable amounts by the respiratory processes within the plant.