"It concerns the fact that certain species of trees do not regularly feed in the soil, but that they are everywhere in their whole root system in symbiosis with a fungus mycelium that serves them like a foster-mother and takes charge for the whole feeding of the tree from the soil... This fungal coat covers the root completely and covers also the entire growing point. It grows as one with the root�s tip and behaves in every respect like an organically linked, peripheral tissue belonging to the root. The whole body is therefore neither tree root nor fungus alone, but a union of two different creatures forming a homogeneous morphological organ similar to the thallus of a lichen and it may be best termed a fungus root, a mycorrhiza..... It (the coat) does not only cover the root tip tightly, but it inserts hyphens through the epidermal cells into the root itself... (though) these hyphens were never traced up to the endodermis..... They do never enter the cells� lumina." (B. FRANK, 1885)
117 years of research into mycorrhiza showed that almost 80 percent of all terrestrial plants can form mycorrhiza-associations. It is distinguished between two basically different types:
- ectotrophic mycorrhizas (or ectomycorrhizas)
- endotrophic mycorrhizas (or endomycorrhizas)
The so-called vesicular-arbuscular mycorrhiza (in short: V/A-mycorrhiza), that has by now shown itself to be the most common type, is among the endomycorrhizas.
Ectomycorrhizas are exactly as described in FRANK�s citation. The fungus forms a coat around the root tip. Its hyphens enter the root�s cortex tissue but not the cells.
Endomycorrhizas form haustoria. A part of the mycelium extends into the cortex cells and between them while another part spreads out into the soil. An entire mycelium coat is usually not formed. The V/A-mycorrhiza is characterized by specific haustoria. Vesicles or tuft-like branched structures (arbuscules) form within the cell lumina. The hyphens of the V/A-mycorrhizal fungi have no partitions. They belong to the zygomycetes. The fungi of ectomycorrhizas and non-V/A-endomycorrhizas are mainly basidiomycetes, more rarely also ascomycetes (the hyphens of these fungi have partitions). Zygomycetes could sometimes also be shown to be part of ectomycorrhizas.
Additional information:
Ectomycorrhizas can be found with a number of tree and shrub species, especially from the Pinaceae, Cupressaceae, Fagaceae, Betulaceae, Salicaceae, Dipterocarpaceae, Myrtaceae, and Caesalpinaceae families. Most mycorrhiza trees live in boreal (cold) and temperate climates, or in tropic habitats poor in nutrients. The fungus serves the tree by supplying water-dissolved minerals as had already been observed by FRANK. Moreover, the fungal coat provides a protection against parasitic fungi of the soil (honey bootlace fungus, Phytophthora (late blight of potatoes), etc.). The protection bases in some cases upon a resistance acquired by the plant and induced by the fungus that can be recognized by the production of defence compounds (phenolic compounds). Mycorrhizal fungi do often secrete phytohormones that have an influence upon root growth. The root system and the fungus mycelium together form a rhizosphere in which living conditions differ from that of the surrounding. The rhizosphere contains, for example, almost always more bacteria than other regions of the soil.
The plant supplies the mycorrhizal fungi with carbohydrates (usually glucose), growth stimulators (volatile terpenes), and vitamins (thiamine or its precursors, pyrimidine or thiazolium, biotin, pantothenic acid, nicotine acid, etc.). Many fungi are heterotrophic regarding vitamins, and are therefore dependent on a supply by the plant. The required spectrum of nutrients differs nevertheless from species to species. Many mycorrhizal fungi can be isolated and cultivated, but they do hardly ever develop fruiting bodies under such conditions.
Factors stimulating the development of fruiting bodies have mostly not been analyzed. Mycorrhizal fungi are usually extremely sensitive against overgrowing of other (saprophytic) fungi. Well-known basidiomycetes like cepe, Amanita, russula, and Lycoperdon belong to the mycorrhiza-formers as well as ascomycetes (like summer truffle). About 5,000 ectomycorrhiza-forming species have been identified world-wide. They are usually not very particular regarding their tree host. Young trees are often associated with other fungi than older ones. Numerous fungi, like the fly agaric, associate with a range of tree species, only few fungus species are found in just one habitat. Among them is Boletus (Boletus elegans) that grows exclusively in larch forests.
The root systems of neighbouring trees of the same or different species are often connected by a shared mycelium (see next section). Ectomycorrhizas are not necessary for seed germination and the early stages of the tree seedlings. Nearly all seedlings that are a few month old live under natural conditions already in association with fungi, though. Controlled experiments showed that seedlings thrive far worse in sterile soil than their counterparts associated with mycorrhizal fungi.
Infected roots differ morphologically from uninfected ones. The cortex cells develop no secondary walls, root hairs and calyptra are missing, and the endodermis is usually impregnated with tannins in order to prevent a further advance of the fungi. The network of hyphens within the cortex is called the HARTIG-net.
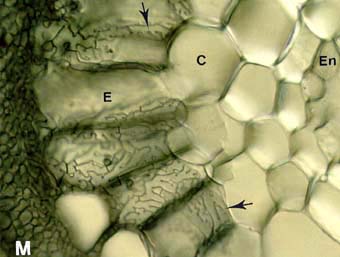
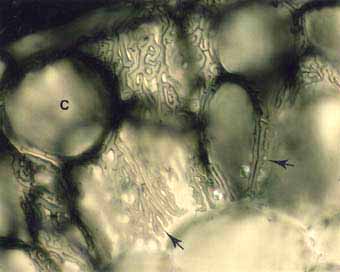
Picture to the left: High magnification view in the Pinus strobus ECM short root. This image shows how Hartig net hyphae (arrows) envelop cortex cells (C). - Picture to the right: Populus tremuloides ECM root cross section showing labyrinthine Hartig net hyphae (arrows) around elongated epidermal cells. This complex hyphal branching pattern is considered to increase the fungal surface area in contact with the root.
In contrast to saprophytic or parasitic fungi, most mycorrhizal fungi secrete no phenoloxidases required for the degradation of lignin and no cellulases necessary for breaking down cellulose. This is true as long as enough glucose is supplied. Cellulase is produced as soon as the supply stops, and the previously symbiotic fungus goes over to a parasitic life style. Two species of mycorrhizal fungi have on the other hand been shown to produce continuously large quantities of cellulase, xylanase, and proteases (no pectinase, though) without causing a damage to the symbiosis. Mycorrhizal fungi prefer slightly acidic soils, pH values between 4.0 and 5.0 are optimal. They cannot exist at a pH higher than 7. mycorrhizas are extremely rare or only rudimentary developed in calcium-rich soils like limestone beech forests. This fact is especially interesting in connection with the rescue actions against the damages caused by the acid rain, since the pH of forest soils is locally drastically increased causing considerable damage of the mycorrhizal fungi. This may lead to secondary damages of the forest trees.
The mentioned more efficient use of the nutrient supply is among the mutual advantages of the mycorrhizas. A larger volume of soil can be tapped, since the fungus mycelium is characterized by a surface far larger than that of the root system. The plant would have to invest much more energy in order to develop an as complex root system without a fungus mycelium than it spends on the fungus in terms of carbohydrates. Carbohydrates are essential for the fungi, though, since they cover their need for nitrogen mainly by the uptake of reduced nitrogen compounds (ammonium- and amino-groups) the fixation of which requires acceptor molecules (carbon skeletons). A lack of carbohydrates inhibits the development of a mycorrhiza, a lack of nitrogen or phosphorus stimulates it.
Fungi secrete protons and acidify the soil. As a countermove(by the development of an electrochemical potential), they take up larger amounts of phosphate and accumulate it within the cells as polyphosphate. A considerable amount of it comes in useful to the plant. Measurements show that especially the phosphate values, not that much also the nitrogen and potassium values are increased in mycorrhizal seedlings compared to the control plants.
The fact that grassland (prairie), that largely lacks mycorrhizal fungi, is very hard to afforest illustrates how important their presence is. Numerous tries failed. Afforesting measures succeeded only after the trees had been inoculated in tree nurseries.
The demarcation of endomycorrhizas from ectomycorrhizas is by no means as sharp and clear as may be assumed from the definition given at the beginning of this topic. Numerous fluid transitions exist and often changes an ectomycorrhiza into an endomycorrhiza. Such cases have been termed ecto-endomycorrhizas. V/A-mycorrhizas are by far the most common types as has been mentioned before. Before going into more detail, two other manifestations, the mycorrhizas of Ericales and that of orchids have to be introduced.
Ericales, and especially species of the Ericaceae-family are in nature always associated with fungi. They are therefore regarded as obligatorily mycotrophic. This does not mean that Ericaceae cannot be cultivated without fungi. Calluna, Vaccinum, azaleas and others grow just as well when cultivated on a purely inorganic substrate as in nature together with fungi. If organic material (peptone or yeast extract, for example) is added to the nutrient medium, however, growth is strongly inhibited. It seems that the roots secrete substances that react with organic material and form toxins. These toxins are deactivated in the presence of fungi. In other words: growth on nutrient-rich soils is optimal in the presence of mycorrhizal fungi. Ericaceae, however, occur almost only on acidic soils extremely poor in nutrients (oligotrophic). The advantage of the obligatory association with fungi allows them to use even these soils efficiently. The fungus attack occurs in a number of species shortly above the vegetative point. Calluna�s primary vegetative point, however, is destroyed by the fungus, and as a consequence secondary vegetative zones are activated causing an increase in the root system�s number of branchings and thus a better penetration of the soil.
It is problematic to afforest Calluna expanses, since the mycorrhizal fungi that forest trees need are lacking. The lack exists due to inhibiting substances, that are secreted by the mycorrhizal fungi that socialize with Calluna. Experiments showed, that extracts from Calluna-humus inhibit the mycelium growth of many fungi. Only a few species like Boletus scaber and Amanita muscaria are resistant. Both species occur everywhere as mycorrhizal fungi where birches begin to penetrate Calluna expanses.
A phenomenon similar to the Ericaceae-mycorrhizas occurs in pinesap (Monotropa), a saprophytic plant devoid of chloroplasts as well as in different species of (green) Pyrolaceae. Monotropa is a root parasite growing on various deciduous trees and conifers. Its root is wrapped in a dense fungus mycelium that penetrates not only the surrounding soil, but also the roots of the host trees thus making an indirect contact between host and parasite. F. FRANCIS and D. J. READ (1984) verified experimentally with the example of a V/A-mycorrhiza that such bridges lead indeed assimilates.
They planted radioactively labelled seedlings of Plantago major together with non-labelled ones of Festuca ovina into a culture container. In a further experiment, Plantago was inoculated with a fungus. The root systems of both species are easily distinguished morphologically. The radioactivity spreads both in the fungus and in Festuca ovina after a short culture period in the container, while the roots of Festuca ovina remained unlabelled despite of the direct physical contact with the labelled Plantago-roots in the fungus-free culture.
Orchidaceae are obligatory mycotrophic just like Ericaceae. Their causes differ, though (H. BURGEFF, 1909, 1936). Orchid seeds are extremely small (0.3 � 14 µm). They have usually no cotyledons which means that a seed can germinate but not develop further than to a few-celled state. Further development is possible only in association with fungi that provide the nutrient substratum. Orchids are accordingly parasites if only in the first phase of their life. Many of them (those with green leaves) change to an autotrophic life during a later stage of development. The fungus becomes superfluous.
The mycorrhizal fungus penetrates the tissue of the young seedling (usually through the suspensor), from where it spreads into the developing root. Shoot and root tuber (if present) are generally not infected.
The endotrophic fungi do usually perish in the course of the plant�s development and their remains are absorbed by the orchids. If this host-specific action does not take place, then the fungus spreads and becomes parasitic. That is why only a small percentage of seedlings develops in many orchid species. The inhibition of the fungus growth is caused by the synthesis of an antagonist that was at first named orchinol, and was later characterized as dehydroxyphenanthrin by E. GÄUMANN and H. KERN (1959). This fungicide has an effect on numerous mycorrhizal and terrestrial fungi. Its synthesis is only induced in the presence of the fungus.


Mature arbuscule of Glomus mosseae with numerous fine branch hyphae. - Arbuscule of Gigaspora margarita with an elongated trunk hypha (T) and tufts of fine branch hyphae (arrows). Note how this arbuscule differs from the Glomus arbuscules (left).
Lobed vesicles of an Acaulospora species in a clover root.
The V/A-mycorrhiza is the most common type of mycorrhiza and has been a focus of research especially during the last years. No plant family, where it has not been detected, exists. It is rare only with Juncaceae, Cyperaceae, Caryophyllaceae, and Brassicaceae. V/A-mycorrhizas are less distinct with plants of humid than with those of dry habitats. Salt meadow plants are often heavily infected. The fungus penetrates the root cortex but never meristems, vascular tissues, chlorophyll-containing or otherwise specialized tissues.
An essential characteristic of the already discussed ectomycorrhizas is the high number of fungus species. In contrast, only 30 morphologically distinguishable forms (species?) of V/A-mycorrhizas have been identified world-wide. The majority of them belongs to the genus Glomus. D. W. MALLOCH, K. A. PIROZYNSKI and P. H. RAVEN pointed out (1980) that V/A-mycorrhizas occur mainly in plant communities rich in species, while ectomycorrhizas are typical for those poor in species. The number of plant species is thus inverse proportional to the number of symbiotic fungi.
Ectomycorrhizas made their first appearance presumably not until the Middle Cretaceous period. Theirs has to be regarded as a progressive form of symbiosis that allowed tree species to survive in temperate and cold climates or on poor soils. The V/A-mycorrhiza is a more primitive symbiosis. It exists since multicellular terrestrial plants exist. Associations between fungi and Rhynia as well as Asteroxylon have been proven by fossil finds from the Devonian era. The most primitive of the contemporary vascular plants (Psilotum and Tmesipteris) are also characterized by V/A-mycorrhizas. It seems reasonable to assume that large trees could not have developed without V/A-mycorrhizas, since they are more wide-spread in wooden plants than in herbs. The mycelium system increases the absorbing surface within the soil many times over as has been mentioned above. Experiments with isotopes showed that 32P spreads in the soil in a given period of time by about 7 mm. The spreading is 7.5 cm in soils interspersed with fungi in the same period of time.
One of the biggest advantages of V/A-mycorrhizas is the additional supplying of plants with phosphate. This statement is safeguarded by numerous experiments. Extremely high contents of phosphate [1 g calcium phosphate per kg soil] inhibit the spreading of the mycelium, though. hemmen.
Plant species (herbs) with a well-developed system of root hairs enter into a symbiosis with fungi only in habitats poor in phosphate. The photosynthetic rate (fixation of carbon dioxide) of associations between plants and fungi is far higher during lack of water than that of specimens without fungi of the same plant species. V/A-mycorrhizas are not very choosy regarding the carbohydrates that are supplied by the plant. They are able to use a wide spectrum of monosaccharides, disaccharides or polysaccharides.
|
|