Zu den Chrysophyta werden Algen gerechnet, die sich durch eine Reihe elektronenmikroskopisch erkennbarer Gemeinsamkeiten auszeichnen. Ein wesentliches Kennzeichen ist eine Falte des Endoplasmatischen Retikulums, die die Plastiden umgibt, sowie die Anordnung der Thylakoide in Dreierstapeln. In der Regel sind zwei unterschiedlich lange Geißeln vorhanden, von denen die längere, nach vorn gerichtete, mit steifen Flimmerhaaren (Mastigonemen) besetzt ist. Zur Pigmentausstattung gehören die Chlorophylle a und c und, mengenmäßig überwiegend, Carotinoide, welche die grüne Chlorophyllfarbe überdecken. Auffallend ist das Fucoxanthin, das den Plastiden vieler Arten eine goldbraune Farbe verleiht.
Der wichtigste Reservestoff ist das Laminaran (ein beta-1,3 -Glucan); doch fast ebenso wichtig sind die vielfach in Vakuolen gespeicherten Öle. Chrysophyta sind meist einzellig, gelegentlich koloniebildend; wenige Arten bilden fadenförmige Thalli aus. In manchen Fällen wichtig. In der Abteilung kommen fast alle morphologischen "Organisationsstufen" (außer parenchymatischen Thalli) vor. Manche Autoren vereinen die Abteilung der Chrysophyta mit der der Phaeophyta zu der gemeinsamen Gruppe der Heterokontophyta. Argumente dagegen werden später zur Diskussion gestellt. Stellt man die
Phaeophyta zunächst zurück, verbleiben drei große Klassen:
Die Unterschiede zwischen ihnen sind beträchtlich, jede stellt eine in sich geschlossene Gruppe dar.
Die Chrysophyceae repräsentieren eine Flagellatenklasse mit etwa 1000 bis 1500 Arten. Man nahm zunächst an, sie seien im Süßwasser häufiger als im Seewasser. Diese Meinung könnte sich ändern, nachdem festgestellt wurde, daß sie einen wesentlichen Anteil am sogenannten Nanoplankton haben. Darunter versteht man planktich lebende Organismen mit Zelldurchmessern in einer Größenordnung von 5 bis 20 µm. Die gängigen Planktonnetze haben Maschenweiten von 40 bis 70 µm, so daß Nanoplankter bei den meisten (älteren) Planktonuntersuchungen nicht mit erfaßt wurden.
Viele Chrysophyceen enthalten Chloroplasten und sind photoautotroph. Manche der Arten bleiben jedoch auf eine Vitaminzufuhr [Thiamin, Biotin (Ochromonas danica) oder Vitamin B12 (Poteriochromonas stipitata)] angewiesen. Der überwiegende Teil von ihnen ernährt sich heterotroph, Phagozytose kommt vor.
Üblicherweise vermehren sich die Chrysophyceen asexuell, bei einigen (z.B. bei Dinobryon) wurde Anisogamie nachgewiesen.
Echte Zellwände sind nicht vorhanden - die Zellform ist vielfach veränderbar -, statt dessen sind die Zellen einiger Gattungen, z.B. Synura, von einem aus silikathaltigen Plättchen bestehenden Panzer umgeben, dessen Synthese elektronenmikroskopisch gut untersucht worden ist. Die Bildung der Plättchen wird im Endoplasmatischen Retikulum initiiert, die fertigen Produkte werden über das intrazelluläre Membransystem an die Zelloberfläche befördert und dort in regelmäßigen Mustern abgelagert.
Andere Chrysophyceen synthetisieren charakteristisch geformte Loricas, welche die Zellen ganz oder teilweise umhüllen. Die Gerüstsubstanz der Lorica von Poteriochromonas stipitata ist Chitin (E. SCHNEPF, W. HERTH; Zellenlehre, Universität Heidelberg). Die Lorica von Dinobryon ist das Gemeinschaftswerk mehrerer Zellen. Die Silikoflagellaten zeichnen sich durch ein intrazelluläres Silikatskelett aus.
Zur Klassifikation: Zu den wichtigen, aber nicht unumstrittenen Kriterien gehört das Vorhandensein und die Zahl der Geißeln. Innerhalb einer jeden der so charakterisierten Gruppen ist entscheidend, ob die Zellen einzeln sind, zu Kolonien zusammentreten, oder welche morphologischen Stufen die Thalli erreichen.

Triceratium (links) und Coscinodiscus, zwei Diatomeengattungen aus der Gruppe der Centrales, aufgenommen in Aufsicht (Valvenansicht), gekammerte Kieselschale (Aufn.: W. KASPRIK).
Die Diatomeen umfassen 6 000 bis 10 000 rezente, meist einzellige Arten. Die Zellen sind diploid. Bei einigen wenigen Arten sind sie zu langen Filamenten vereint. Die Angaben über die Zahl fossiler Arten schwanken zwischen 40 000 und 100 000. Das auffallende Merkmal dieses Taxons ist eine silikathaltige, dauerhafte Schale (Frustel). Ihretwegen sind sie als Fossilien gut erhalten (Kieselgur). Ein Kubikzentimeter Kieselgur enthält etwa 4,6 Millionen Schalen. In Ablagerungen erreichen Kieselgurschichten Mächtigkeiten von mehreren 100 Metern.
Die Systematik der Diatomeen beruht ausschließlich auf der Auswertung von Unterschieden in der Architektur der Schalen (Größe, Symmetrie, Skulptierung u.a.). Doch wie im Abschnitt "Bewertung morphologisher Merkmale..." dargelegt, ist das ein höchst unbefriedigendes Unterfangen. In kaum einer anderen Organismengruppe ist der Streit, ob zwei Formen einer Art oder zwei Arten zuzuordnen sind, so groß wie hier.
Die meisten Artbeschreibungen beziehen sich auf kleine, nicht repräsentative Stichproben. Nur selten weiß man Genaueres über die Variabilität der einzelnen Schalenparameter, und erst bei einigen wenigen Arten hat man zeigen können, daß Temperatur, pH-Wert, Salinität sowie Phosphat- und Silikatgehalt des Mediums die Form der Schalen merklich beeinflussen. Durch lichtmikroskopische Beobachtung (wie bisher üblich) sind Strukturdetails vielfach nur schwer erkennbar. Deshalb hat der Einsatz der Rasterelektronenmikroskopie die Diagnostik der Diatomeen entscheidend verbessert. Nur wenige Arten konnten in Kultur genommen werden, und nur sehr selten sind die einzelnen Stadien des Lebenszyklus voll erfaßt
Bau der Schale (Frustel): Diatomeenschalen nennt man Frusteln. Sie bestehen aus zwei unterschiedlich großen Teilen, die schachtelförmig oder wie Deckel und Boden einer Petrischale aufeinandergesetzt erscheinen. Der Deckel (die größere, obere Hälfte) wird als Epitheka, der Boden (die kleinere, untere Hälfte) als Hypotheka bezeichnet. Die in Aufsicht erkennbaren Flächen nennt man Valven. In Seitenansicht sieht man den Gürtel (das Gürtelband) und spricht daher auch von Gürtelansicht. Bei jeder Zellteilung wird die Hypotheka ersetzt. Bleiben nach der Teilung beide Hälften erhalten, entstehen in der einen Nachkommenschaftslinie ständig kleiner werdende Zellen. Die Reduktion der Größe kann je nach Art 30 bis 50 Prozent betragen.
Viele Arten bilden unter ungünstigen Lebensbedingungen Dauerstadien (Zysten, Ruhestadien) mit verstärkter Wand aus.
Die Bacillariophyceae können in zwei klar voneinander getrennte Ordnungen untergliedert werden, die sich in der Symmetrie der Schale und in ihrem Fortpflanzungsverhalten voneinander unterscheiden. Die Pennales zeichnen sich durch bilaterale Symmetrie und geißellose Isogameten aus, die Centrales durch Radiärsymmetrie. Ihre männlichen Gameten sind aktiv beweglich begeißelt, wenn auch eine der beiden Geißeln fehlt und das Axonem vereinfacht sein kann, die weiblichen unbegeißelt (Oogamie).
Die Schale der Diatomeen besteht primär aus pektinartigen Polysacchariden und Proteinen; Cellulose wurde nicht nachgewiesen. Siliciumoxyd wird in speziellen membranumschlossenen Vesikeln (den SDVs = silica depositing vesicles) unterhalb der künftigen Zelloberfläche im Cytoplasma deponiert und bildet komplex gebaute Strukturen aus. Die Schalen sind artspezifisch durch regelmäßig angeordnete Poren oder Rillen durchbrochen, so daß ein Kontakt zwischen Zellinhalt und Umgebung gewahrt bleibt; an anderen Stellen sind sie durch Vorsprünge, Auswüchse, Wülste usw. verstärkt.
Bei vielen Arten sind deutlich erkennbare Schwebefortsätze ausgebildet (in Fossilien oft nicht erhalten). Bei einigen Arten (Gattungen), so z.B. bei Thalassiosira fluviatilis und mehreren Cyclotella-Arten, bestehen sie aus Chitin. Der Auftrieb wird auch durch niedriges spezifisches Gewicht der Zellen - viele enthalten große Vakuolen und Öl als Reservestoff - erhöht. Diatomeen scheiden oft Gallerten aus, die in sich homogen erscheinen, molekular aber sehr heterogen strukturiert sein können. Bei Pennales werden kohlenhydrathaltige Gallerten oft nur an den Polen abgesondert. Sie dienen u.a. dem Zusammenhalt von Zellen, und je nach Art entstehen sternförmige, fächerförmige oder kettenförmige (zick-zackförmige) Aggregate. Die Gallerten dienen aber auch der Anheftung (Adhäsion) der Zellen an lebende oder tote Unterlagen.
Die Valven vieler Pennales sind durch unterschiedlich gebaute, längs verlaufende Kanäle (Raphen) durchbrochen. Raphenhaltige Zellen bewegen sich gleitend. Welcher molekulare Mechanismus dem zugrunde liegt, ist unklar. Fest steht, daß die Geschwindigkeit mit bis zu 20 mm/sec. sehr hoch ist und daß bei der Bewegung eine Bewegungsgallerte ausgeschieden wird, die in der Raphe entlanggleitet und Schleimspur hinterläßt.
Vorkommen und ökologische Bedeutung: Die meisten Centrales-Arten leben planktisch. Pennales sind planktisch oder benthisch. Auf sie gehen die braunen Überzüge (Fucoxanthin) vieler grüner Wasserpflanzen (epiphytische Lebensweise) und die Braunfärbung des Watts zurück . Die Centrales bilden die Hauptmasse des Phytoplanktons der Ozeane. Auffallend ist die hohe Produktion in kalten Gewässern: in der Antarktis, in Aufquellgebieten vor der Küste Südwestafrikas und an den Westküsten Südamerikas und Kaliforniens.
Man hat die Primärproduktion auf 200 - 400 g Biomasse/Quadratmeter/Jahr errechnet. Zum Vergleich: auf Getreide- oder Kartoffelfeldern liegen die Werte bei 500 - 1000 g Biomasse/Quadratmeter/Jahr.
Im Verlauf des Jahres (Jahreszyklus) erscheint die Hauptmasse an marinen Diatomeen in der gemäßigten Klimazone (z.B. in Nord- und Mitteleuropa) unmittelbar im Anschluß an das Schmelzen des Eises. Zu dieser Zeit (Februar/Anfang März) sind die Gewässer nährstoffreich (reich an Phosphaten, Nitraten, Sulfaten u.a.) und für sie wichtiges Silikat ist verfügbar; mit zunehmender Tageslänge verbessern sich die Voraussetzungen zur Photosynthese. Die Diatomeen haben unter diesen Bedingungen eine ökologische Nische erobert. Ein zweites, meist deutlich niedrigeres Maximum erreicht die Populationsdichte gegen Ende des Sommers (August bis Oktober).
J. C. GALlAGHER zeigte 1980, daß sich Frühjahrs- und Herbstpopulation der Art Skeletonema constatum (Centrales) in der Narragansett-Bay (Rhode Island, Ostküste USA) morphologisch zwar gleichen, genetisch sich aber voneinander unterscheiden. Er isolierte mehrere hundert Zellen aus beiden Populationen und kultivierte die sich durch Teilung daraus bildenden Nachkommenschaftsklone unter Laborbedingungen getrennt weiter. Als Maß für die genetische Variabilität analysierte er das Alloenzymmuster fünf verschiedener Enzyme in jedem der Klone. In jeder Population fand sich eine beachtliche Variabilität, keiner der Klone war, für sich alleine genommen, für die eine oder die andere Population repräsentativ, aber im Durchschnitt ergaben sich signifikante Häufigkeitsunterschiede der Alloenzyme zwischen beiden Populationen. Es sieht demnach so aus, als sei der Genfluß zwischen beiden unterbrochen oder zumindest stark eingeschränkt.
Fortpflanzung: Normalerweise vermehren sich die Diatomeen asexuell. Nach Erreichen einer Minimalgröße kann es - nach erfolgter Meiose - zur Gametenbildung kommen. Das Verschmelzen zweier Gameten führt zu einer, hier Auxospore genannten Zygote. Die Auxosporen nehmen an Größe zu und verdrängen die Schalen, innerhalb derer sie entstanden sind. Sie haben nach der Freisetzung zunächst keine typische Schale, doch folgt die Bildung arttypischer erheblich größerer Theken in Verbindung mit speziellen Mitose- und Plasmolysevorgängen unmittelbar im Anschluß an das Schlüpfen. Die fertigen Zellen werden Erstlingszellen genannt.
Asexuelle und sexuelle Fortpflanzung bei Diatomeen. Die Schalengröße nimmt bei asexueller Vermehrung in einer der Tochterzellinien sukzessive ab. Nach Erreichen einer minimalen Größe (ca. 30 % der maximalen Größe) durchlaufen die Zellen eine Meiose. Etwa die Hälfte solcher Zellen (Gametophyten) produziert Spermien, die andere Eier (Oogonien). Nach dem Verschmelzen der Gameten entsteht eine Auxospore (eine Zygote), die sich zu einer mit einer silikathaltigen Schale versehenen Zelle differenziert. Der Kreis ist damit geschlossen (Nach P. H. RAVEN, R. F. EVERT, H. CURTIS, 1981).
Centrales und Pennales zeichnen sich durch folgende Besonderheiten aus: Viele (nicht alle) Arten der Centrales bilden begeißelte männliche Gameten. Im Unterschied zu den sonst üblichen Eukaryotengeißeln ("9 + 2") fehlen hier die beiden zentral gelegenen Mikrotubuli. Die Gameten sind geißellos und in der Regel größer als die männlichen. Bei Pennales sind nie begeißelte Stadien gesehen worden. Nach Annäherung zweier paarungsbereiter (kleiner) Zellen bilden sie eine gemeinsame Gallerthülle (Kopulationsgallerte) aus. Anschließend durchlaufen beide Zellen die Meiose; woraufhin häufig zwei Gameten entstehen (zwei der Kerne gehen zugrunde). Die Theken weichen auseinander, und die wandlosen Gameten können mit denen der anderen Zelle fusionieren. Neben dieser, als Allogamie (Fremdbefruchtung) zu bezeichnenden Alternative, findet man auch Autogamie, die auf dem Verschmelzen zweier Gameten aus der gleichen Zelle beruht sowie Apomixis, bei der der Prozeß der Gametenbildung eingeleitet wird, Meiose und Gametenfusion jedoch unterbleiben.
Systematische Untergliederung der beiden Ordnungen.
Pennales: Die Untergliederung in Unterordnungen erfolgt aufgrund der An- oder Abwesenheit und der Ausgestaltung der Raphe.
Biraphidinae: Raphe auf beiden Valven.
Monoraphidinae: Raphe nur auf einer Valve.
Raphidioidineae: Raphe nur rudimentär ausgebildet, z.B. nur an den beiden Polen der Zelle.
Araphidinae: ohne Raphe.
Centrales: Die Einteilung in Unterordnungen beruht auf der Form der Zellen in Aufsicht (Valvenansicht).
Coscinodiscineae: Valven rund (flach oder konvex), meist ohne auffallende Fortsätze. Der Durchmesser der Frusteln ist größer als die Höhe.
Rhizosoleniineae: Frusteln langgestreckt, zylindrisch.
Biddulphiineae: Valven bipolar oder multipolar gebaut, an den Ecken Fortsätze oder Verdikkungen.
Die Xanthophyceae (= Tribophyceae) ähneln den Chrysophyceae, Fucoxanthin fehlt jedoch, Chlorophyll c kommt bei einigen Arten vor. Sie ähneln aber auch den Chlorophyta, es fehlt ihnen jedoch das Chlorophyll b und die Stärke als Reservestoff. Die Artenzahl beträgt etwa 400. Keine Art ist wirklich häufig, keine ist an Wasserblüten beteiligt. Die meisten leben im Süßwasser oder im feuchten Boden, einige wenige im Brack- und im Seewasser.
Sie vermehren sich asexuell und sexuell (durch Oogamie). Neben einzelligen Arten gibt es Arten, deren Thallus aus mehrzelligen unverzweigten oder verzweigten Fäden besteht (z.B. Tribonema) oder siphonal organisiert ist. Unter siphonal versteht man die Ausbildung querwandloser, schlauchartiger, teils verzweigter, teils mit Rhizoiden versehener, vielkerniger Thalli. Typische Beispiele hierfür sind Vaucheria und Botrychium.
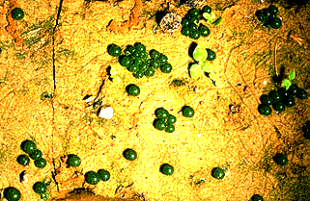
Botrychium granulatum. Die grünen Kugeln sind mit rhizoidartigen Fortsätzen im trockenfallenden Schlamm verankert. Bei Benetzung entlassen sie Zoosporen
(Aufn.: W. KASPRIK).
Die meisten Arten sind - zumindest in bestimmten Lebensphasen - begeißelt. Neben zweigeißligen kommen vielgeißlige Arten (oder Stadien) vor. Die vegetative Vermehrung siphonaler und vielzelliger Arten erfolgt durch Ausbildung begeißelter, zellwandloser Sporen (Zoosporen). Einige der einzelligen Arten sind stets zellwandlos, in anderen Fällen sind die Zellen zeitweilig oder ständig amöboid. Die Untergliederung der Xanthophyceae erfolgt aufgrund der Organisationshöhe ihrer Thalli. Die wichtigsten Ordnungen und ihre vegetativen Merkmale sind:
Heterochloridales: zweigeißlige, zellwandlose Einzeller; einige Arten leben zeitweilig amöboid.
Rhizochloridales: Arten mit vorwiegend amöboider Lebensweise.
Heterogloeales: coccale Formen (= rund, unbeweglich), einzeln oder zu Kolonien zusammengeschlossen, einige in eine Gallerthülle eingebettet. Verbreitung durch Zoosporen.
Tribonematales: Thalli aus unverzweigten oder verzweigten Filamenten, zweigeißlige Zoosporen.
Vaucheriales: vielkernige, siphonale Thalli.
Bei der vegetativen Vermehrung von Vaucheria entstehen vielgeißlige Schwärmer, die, nachdem sich der terminale Abschnitt durch eine Querwand abgegliedert hat, an den Schlauchenden freigesetzt werden. Die Schwärmer von Vaucheria sind nicht mit Spermatozoiden zu verwechseln. Jene entstehen bei sexueller Fortpflanzung, sie sind viel kleiner als die Schwärmer und zweigeißlig.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de