

"A rare swamp plant."
Characteristics of habitats are the abiotic and biotic factors that influence a plant. Among the abiotic factors are chemical and physical values like light, climate, soil composition, soil consistency, exposition to the sun, etc. Biotic factors are, for example, the influences that a species exerts on other species or on itself (competition, coexistence, etc.).
In 1840, J. v. LIEBIG announced the "Concept of the Minimum". It says that the element that is present in the smallest amount restricts plant growth. The practical effects of this observation are well-known, mentioning artificial fertilisation should be enough. In most agricultural areas, the lack of nitrogen and phosphate compounds limits plant growth. Trace elements, too, can be limiting under natural conditions. Among them is Zn that plants living in sunny habitats need more than plants living in the shadow. Zn is therefore more likely to be a growth limiting factor in sunny than in shadowy habitats.
V. E. SHELFORD enhanced LIEBIG’s concept in 1913. He developed the "Concept of Tolerance". It states that plants are always influenced by a whole bunch of factors and that the plant has a tolerance for each of these factors that can be measured. The by far most important factors are temperature, water, and light as well as a combination of these three.
The most important ecosystems of the earth (map of the earth’s vegetation). The coding is based on the system of classification developed by the UNESCO. It was assembled by ELAINE MATTHEWS, NASA Goddard Institute for Space Studies, New York.
1. (light green) well-tempered rain forests, 2. evergreen boreal coniferous woods, 3. (green) tropical rain forests, 4. (dark green) summer-green deciduous forests, 5. (yellowish orange) tropical dry forests, 6. (orange) dry forests of temperate areas, 7. (light violet) thorny and succulent vegetation, 8. (violet) semi-deserts, 9. (dark violet) xeromorphic vegetation 10. (light blue) dry steppe, 11. dry savannah, 12. (blue) moist savannah, 12, 13. (dark blue) pasture, steppe, 14. (brown) deserts, 15. (white) tundra, 16. (grey) ice
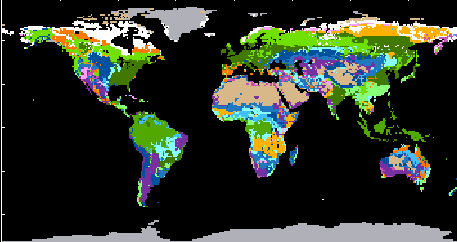
The earth’s vegetation is subdivided into a number of vegetation zones. Each of them is characterised by certain temperatures and humidities, though it has to be considered that daily and seasonal fluctuations are often more important than average values.
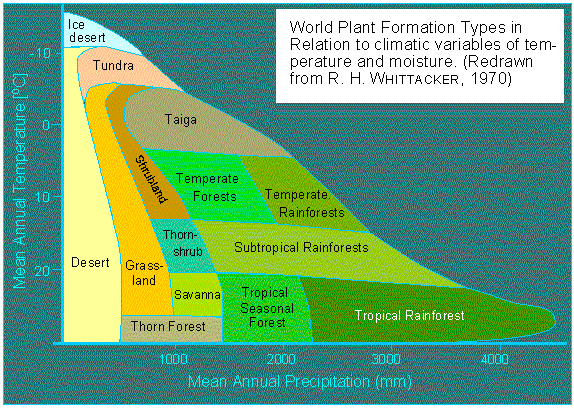
Illustration: Vegetation Zones. Classification of the climax societies according to the average yearly temperature and the amount of rain (according to R. H. WHITTACKER, 1970).
The zones of altitude in the vegetation of mountains has similar causes. The tree line in the south of the Alps, for example, is – caused by the higher average temperatures - higher than that of the north side.
Profile of the Vegetation of the middle east Alps. The limits of altitude are highest in the continental central Alps (according to H. MAYER, 1974).
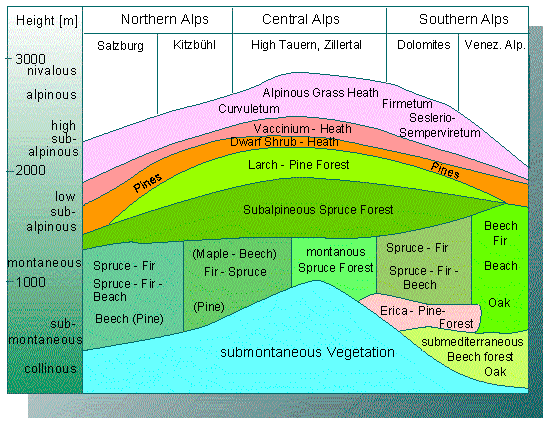
Soil conditions, local climates, and biotic factors control the distribution of single species in agricultural areas, while the overall climate is the reason for the existence of climatic and vegetational zones and thus also of agricultural areas. In its simplest case, the distribution of a species mirrors the geological conditions. This is illustrated very well by the different vegetations of the lime stone alps (Kalkalpen) and the Urgesteinsalpen.
The geographic distribution of each plant species can be determined. Maps of plant distribution drawn from these data do often show that closely related plants are characterised by their different geographic distribution.
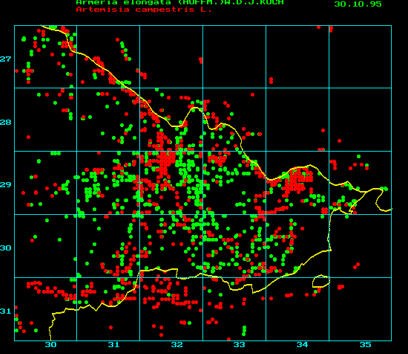
Example of a map of vegetation (regional office for floristic mapping of the administrative district Lüchow-Dannenberg. The project is led by H. W. KALLEN). In order to map and store the data, certain software like Flora-Pro or similar programs is used. The maps of vegetation gain considerably in value by summing up different sets of data in one map as, for example, by comparing the areas of distribution of different species. A linear distribution points either towards a river or a street. Armerica elongata in the diagram above (green) occurs more often in the centre of the administrative district and is distributed along the streets.
In 1988, H. HÄUPLER and P. SCHÖNFELDER published the maps of vegetation of (former Western) Germany as a book. Newer studies are collected in the ‘Project Data Base of Vascular Plants (Central Office for the Floristic Mapping of Germany)’ (http://www.biologie.uni-regensburg.de/Botanik/Florkart/") (see also an example of a map). Its aims are:
To capture and collect the data of the distribution of ferns and flowering plants in the single state and the looking after of regional floristic capturing of data by instructions and EDV, the co-ordination of the single projects and the standardisation of the methods, and the building and looking after of a central data base with the aims:
- to evaluate the data with regard to natural conservation and scientific analysis, like lists of species and biodiversity,
- to observe the situation of the vegetation and its development; to determine ecological groups of species and types of areas; to develop a concept for its monitoring.
- to draw maps of the floristic distribution.
It is distinguished between closed areas, i.e. homogeneous areas of distribution and mosaic-like distribution. Such patterns point at an earlier unified area of distribution. Usually, it cannot be ascertained, whether the separation has physical causes (like the continental drift or ice) or whether they show the beginning dying out of the respective species. Species that occur world-wide are called cosmopolitan, while those occurring only in certain areas like, for example, on certain islands, are called endemic. 90% of all vascular plants living on Hawaii, for example, are endemic. This high percentage is a proof of the very old age of these islands and the long period during which the plants could develop in isolation.
The distribution of species tells something about their potential of distribution. E. MAYR collected several criteria in 1965 that determine the distribution of plants and relate their potential of distribution to that of other organisms:
The potential of distribution of plants is usually larger than that of higher vertebrates. They cross large waters more easily.
Birds and mammals have an easier time establishing themselves in a new surrounding than plants have. Birds and mammals are thus better in colonising.
Within a given climatic zone, plants are more distributed than vertebrates. Vertebrates, in contrast, have an advantage when changing from one climatic zone to another.
The turnover due to colonising and extinction on islands is higher with birds and mammals than with plants. This cannot be extrapolated for all groups of animals. Many insects and lower vertebrates display patterns of distribution that resemble plants more than higher vertebrates.
Soil conditions are decisive for the thriving of plants besides the mentioned climatic factors and water. The soil properties are influenced both by the climate and the organisms. The upper soil layers are therefore always a mixture of inorganic and organic compounds. Soil science distinguishes between horizontal layers termed A-, B-, and C-horizons. The A-horizon covers the uppermost layer that consists mostly of dead plant and animal material. The particle size decreases with increasing depth. Putrefaction and the production of humus take mainly part in the A-horizon. Mineralisation occurs in the B-horizon.
The inorganic components released from organic substances by micro-organisms add to the already present minerals. The C-horizon, finally, consists almost entirely from minerals. The horizons and the single layers within them can in soil profiles often be recognised by their characteristic colour and consistency. The differ in thickness depending on climatic zone, topographic situation and vegetation. Soils that are covered by grass have a very fast humification while mineralisation is slow. Large quantities of organic substances enter the soil as gramineae are usually rather shor-lived, the plant material decays quickly and a lot of humus is produced. This is not true for sandy soils, as all organic material is immediately washed out. In moors, in contrast, a thick layer of humus accumulates and mineralisation does not take place. The humus production of forests is very slow, while mineralisation occurs very fast. Prairie soil contains 600 tons of humus per hectare, forests soil only 50. The almost completely lacking humus production of tropical rain forests and the accompanying fast mineralisation causes the development of laterit and Ortsstein in shallow depths.
These few remarks show, how large the part of plants is in the production and quality of soils and that misuse and false expectations can cause irreversible damage.
Changing properties of habitats cause the extinction of certain species or communities of species and their replacement by others. The influence of humans is especially grave as their methods of cultivation like cutting down of forests, drying moors, river straitening, etc. change the vegetation basically. The addition of a single components is often enough to cause the death of plant species. Many species living on poor, lime-containing grassland are very sensitive towards fertilisation. Among them are many orchids as well as Koeleria gracilis, Pulsatilla vulgaris, Potentilla arenaria, Trifolium scabrum, and others.
Plants can often be used as indicators of certain environmental conditions, but the results should be analysed with care, as the presence or absence of a certain plant species does usually not allow safe conclusions as to the type of local conditions or disruptions.
Large species are usually better indicators than small ones as they have a more stable biomass that is more resistant towards small and short-lived fluctuations of the system. Single-celled algae, in contrast, are ver bad indicators of a water’s quality. The analysis of extensive data showed that certain species may disappear as a result of pollution, but they do very often reappear after having adapted to the new conditions. Whether a species occurs in a given year depends among other factors on its starting conditions. In one year a certain species may be dominant, in another another even though nothing has changed in the physical or chemical parameters of the water. A whole plant community is therefore usually a better indicator than a single species.
Plants may compensate unfavourable conditions considerably. Different habitat conditions cause different phenotypes, different genotypes are selected, the metabolism is changed (isoenzymes), alternative metabolic pathways are taken, and the anatomy is often altered (sun and shade leaves). As has been discussed before, the development of each plant is controlled by different extern factors, especially by light and temperature.
As fluctuations of temperature are lesser in water than on earth, the tolerance of aquatic organisms towards temperature is far lower.
A rising light intensity causes both in terrestrial and aquatic plants an increased photosynthetic activity. The optimum is reached while the values of light saturation are still low. This is especially true for species that have adapted to shadowed biotops or aquatic lifestyles. The highest activity of populations of single-celled algae is thus not measured directly at the water surface, but several decimetres below. Species, that have adapted to sunny habitats and have a C4 metabolism of photosynthesis do even at light saturation not reach the maximal activity.
Not only the absolute amount of rain and the soil moisture are important when analysing the humidity. Important, too, are the seasonal distribution and the balance between rainfall and evaporation. A plant wilts as soon as its rate of transpiration is higher than its rate of water absorption. The growth rate of plants is usually directly proportional to their rate of transpiration, i.e. transpiration and primary production have to be coupled. M. L. ROSENZWEIG (University of Pennsylvania, 1968) described this with the following equation:
log PnN = (1,66 +/- 0,27) log AE - (1,66 x 0,07)
Pn is the rate of net production above the soil (in gram) per square metre, while AE is the yearly transpiration in millimetre. When knowing the latitude, the average monthly temperature and the rainfall, AE can be found in tables, i.e. the net primary production can be calculated. The equation can be used for every ecosystem that is outbalanced, it is less well-suited for developing ecosystems.
|